

Structural insight into dynamic bypass of the major cisplatin-DNA adduct by Y-family polymerase Dpo4
- PMID: 20512114
- PMCID: PMC2892363
- DOI: 10.1038/emboj.2010.101
Structural insight into dynamic bypass of the major cisplatin-DNA adduct by Y-family polymerase Dpo4
Abstract
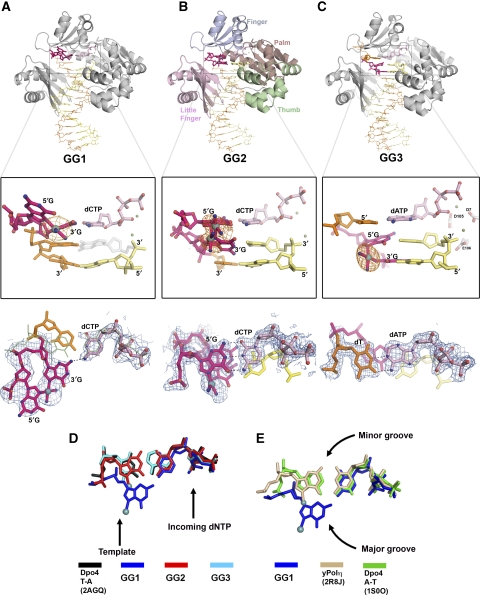
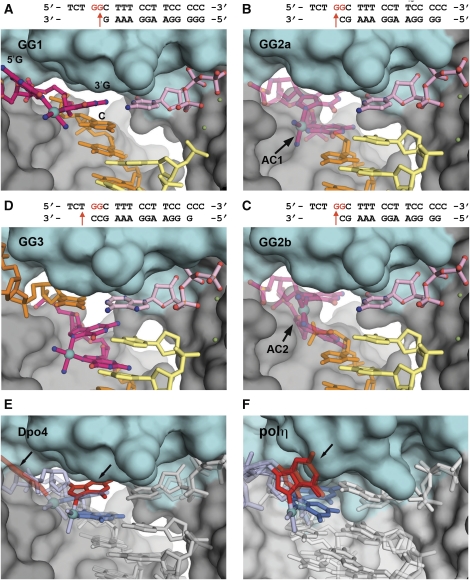
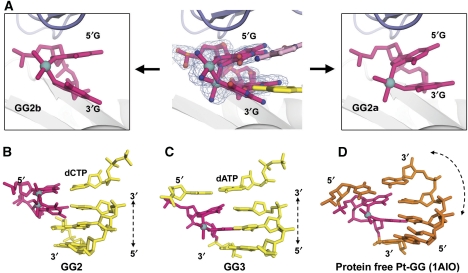
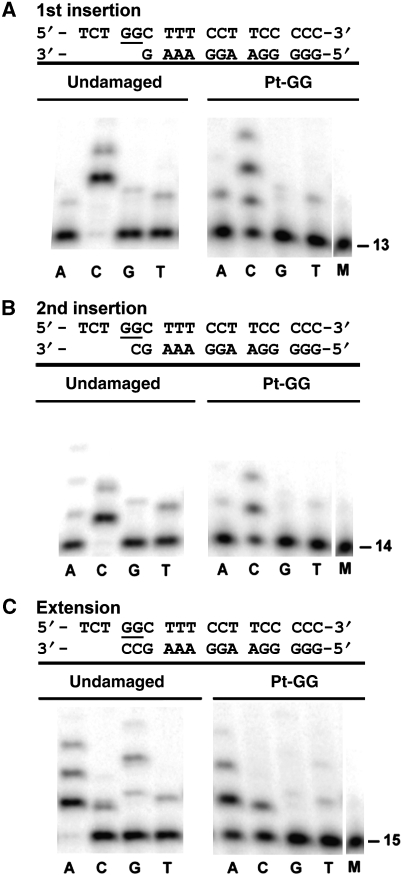
Y-family DNA polymerases bypass Pt-GG, the cisplatin-DNA double-base lesion, contributing to the cisplatin resistance in tumour cells. To reveal the mechanism, we determined three structures of the Y-family DNA polymerase, Dpo4, in complex with Pt-GG DNA. The crystallographic snapshots show three stages of lesion bypass: the nucleotide insertions opposite the 3'G (first insertion) and 5'G (second insertion) of Pt-GG, and the primer extension beyond the lesion site. We observed a dynamic process, in which the lesion was converted from an open and angular conformation at the first insertion to a depressed and nearly parallel conformation at the subsequent reaction stages to fit into the active site of Dpo4. The DNA translocation-coupled conformational change may account for additional inhibition on the second insertion reaction. The structures illustrate that Pt-GG disturbs the replicating base pair in the active site, which reduces the catalytic efficiency and fidelity. The in vivo relevance of Dpo4-mediated Pt-GG bypass was addressed by a dpo-4 knockout strain of Sulfolobus solfataricus, which exhibits enhanced sensitivity to cisplatin and proteomic alterations consistent with genomic stress.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

Similar articles
-
Structural and functional analysis of Sulfolobus solfataricus Y-family DNA polymerase Dpo4-catalyzed bypass of the malondialdehyde-deoxyguanosine adduct.Biochemistry. 2009 Aug 4;48(30):7079-88. doi: 10.1021/bi9003588. Biochemistry. 2009. PMID: 19492857 Free PMC article.
-
Mechanistic studies of the bypass of a bulky single-base lesion catalyzed by a Y-family DNA polymerase.J Biol Chem. 2009 Mar 6;284(10):6379-88. doi: 10.1074/jbc.M808161200. Epub 2009 Jan 5. J Biol Chem. 2009. PMID: 19124465 Free PMC article.
-
Structure-function relationships in miscoding by Sulfolobus solfataricus DNA polymerase Dpo4: guanine N2,N2-dimethyl substitution produces inactive and miscoding polymerase complexes.J Biol Chem. 2009 Jun 26;284(26):17687-99. doi: 10.1074/jbc.M109014274. J Biol Chem. 2009. PMID: 19542237 Free PMC article.
-
The chromosome replication machinery of the archaeon Sulfolobus solfataricus.J Biol Chem. 2006 Jun 2;281(22):15029-32. doi: 10.1074/jbc.R500029200. Epub 2006 Feb 8. J Biol Chem. 2006. PMID: 16467299 Review.
-
Recent insight into the kinetic mechanisms and conformational dynamics of Y-Family DNA polymerases.Biochemistry. 2014 May 6;53(17):2804-14. doi: 10.1021/bi5000405. Epub 2014 Apr 23. Biochemistry. 2014. PMID: 24716482 Free PMC article. Review.
Cited by
-
Promutagenic bypass of 7,8-dihydro-8-oxoadenine by translesion synthesis DNA polymerase Dpo4.Biochem J. 2020 Aug 14;477(15):2859-2871. doi: 10.1042/BCJ20200449. Biochem J. 2020. PMID: 32686822 Free PMC article.
-
Beyond the Lesion: Back to High Fidelity DNA Synthesis.Front Mol Biosci. 2022 Jan 5;8:811540. doi: 10.3389/fmolb.2021.811540. eCollection 2021. Front Mol Biosci. 2022. PMID: 35071328 Free PMC article. Review.
-
Roles of the four DNA polymerases of the crenarchaeon Sulfolobus solfataricus and accessory proteins in DNA replication.J Biol Chem. 2011 Sep 9;286(36):31180-93. doi: 10.1074/jbc.M111.258038. Epub 2011 Jul 22. J Biol Chem. 2011. PMID: 21784862 Free PMC article.
-
Roles of the Y-family DNA polymerase Dbh in accurate replication of the Sulfolobus genome at high temperature.DNA Repair (Amst). 2012 Apr 1;11(4):391-400. doi: 10.1016/j.dnarep.2012.01.005. Epub 2012 Feb 4. DNA Repair (Amst). 2012. PMID: 22305938 Free PMC article.
-
Modulation of Kinase Activities In Vitro by Hepatitis C Virus Protease NS3/NS4A Mediated-Cleavage of Key Immune Modulator Kinases.Cells. 2023 Jan 25;12(3):406. doi: 10.3390/cells12030406. Cells. 2023. PMID: 36766748 Free PMC article.
References
-
- Albertella MR, Green CM, Lehmann AR, O'Connor MJ (2005) A role for polymerase eta in the cellular tolerance to cisplatin-induced damage. Cancer Res 65: 9799–9806 - PubMed
-
- Allen MB (1959) Studies with Cyanidium caldarium, an anomalously pigmented chlorophyte. Arch Mikrobiol 32: 270–277 - PubMed
-
- Alt A, Lammens K, Chiocchini C, Lammens A, Pieck JC, Kuch D, Hopfner KP, Carell T (2007) Bypass of DNA lesions generated during anticancer treatment with cisplatin by DNA polymerase eta. Science 318: 967–970 - PubMed
-
- Bassett E, Vaisman A, Havener JM, Masutani C, Hanaoka F, Chaney SG (2003) Efficiency of extension of mismatched primer termini across from cisplatin and oxaliplatin adducts by human DNA polymerases beta and eta in vitro. Biochemistry 42: 14197–14206 - PubMed
-
- Bassett E, Vaisman A, Tropea KA, McCall CM, Masutani C, Hanaoka F, Chaney SG (2002) Frameshifts and deletions during in vitro translesion synthesis past Pt-DNA adducts by DNA polymerases beta and eta. DNA Repair (Amst) 1: 1003–1016 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
