

Recovery of motoneuron and locomotor function after spinal cord injury depends on constitutive activity in 5-HT2C receptors
- PMID: 20512126
- PMCID: PMC3107820
- DOI: 10.1038/nm.2160
Recovery of motoneuron and locomotor function after spinal cord injury depends on constitutive activity in 5-HT2C receptors
Abstract
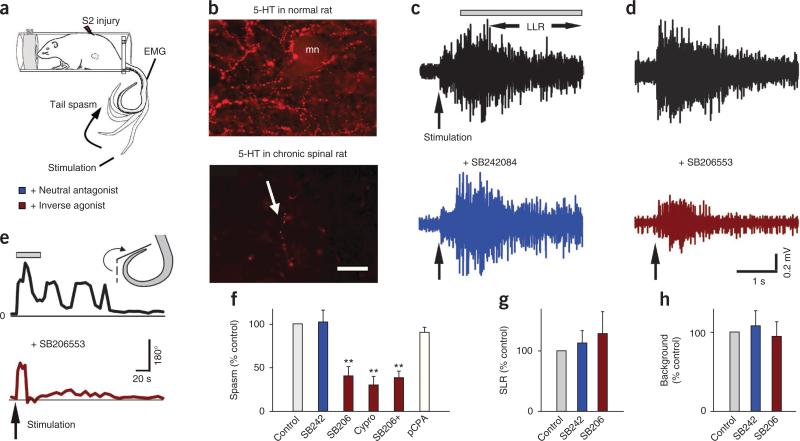
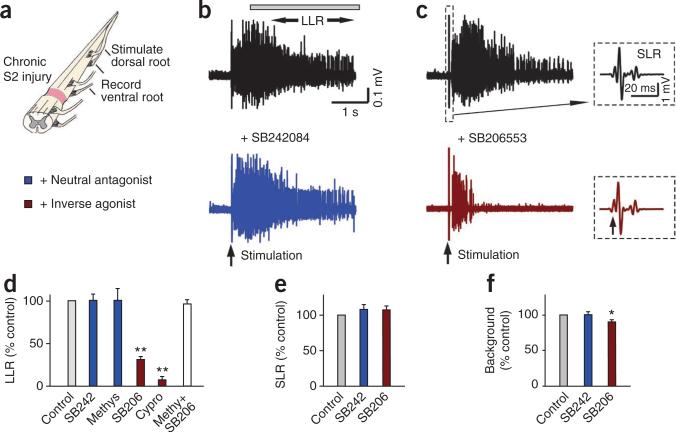
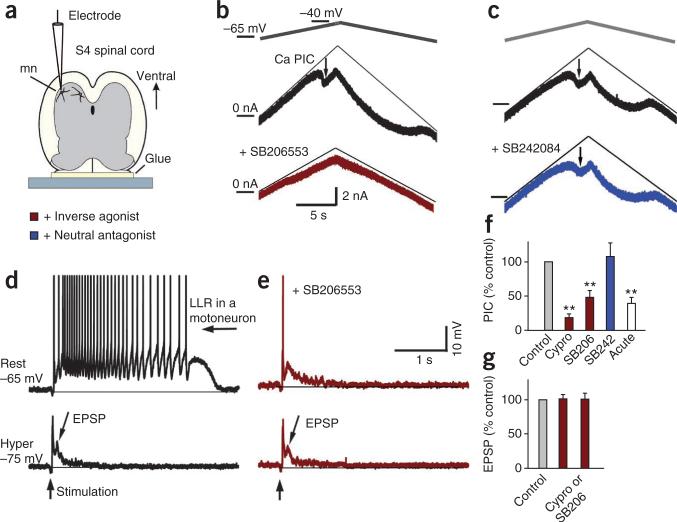
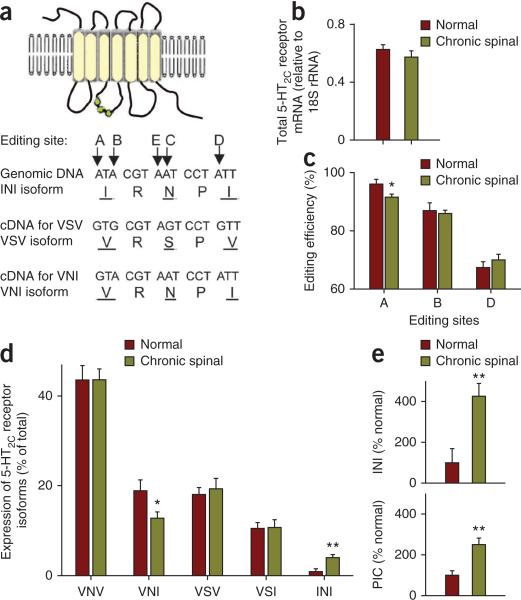
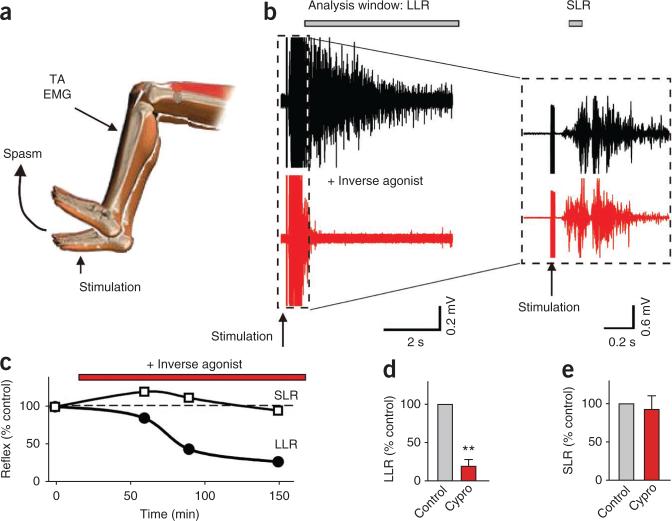
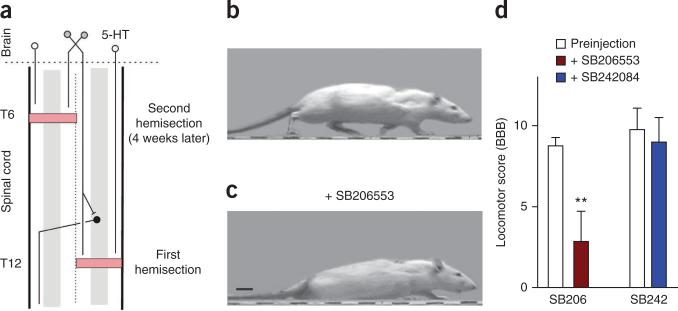
Muscle paralysis after spinal cord injury is partly caused by a loss of brainstem-derived serotonin (5-HT), which normally maintains motoneuron excitability by regulating crucial persistent calcium currents. Here we examine how over time motoneurons compensate for lost 5-HT to regain excitability. We find that, months after a spinal transection in rats, changes in post-transcriptional editing of 5-HT2C receptor mRNA lead to increased expression of 5-HT2C receptor isoforms that are spontaneously active (constitutively active) without 5-HT. Such constitutive receptor activity restores large persistent calcium currents in motoneurons in the absence of 5-HT. We show that this helps motoneurons recover their ability to produce sustained muscle contractions and ultimately enables recovery of motor functions such as locomotion. However, without regulation from the brain, these sustained contractions can also cause debilitating muscle spasms. Accordingly, blocking constitutively active 5-HT2C receptors with SB206553 or cyproheptadine, in both rats and humans, largely eliminates these calcium currents and muscle spasms, providing a new rationale for antispastic drug therapy.
Figures
Comment in
-
Behind the paper: muzzling muscle spasticity.Nat Med. 2010 Jun;16(6):637. doi: 10.1038/nm.2169. Epub 2010 May 30. Nat Med. 2010. PMID: 20512125 No abstract available.
-
Spinal cord injury: the two faces of spontaneous serotonin signalling.Nat Rev Drug Discov. 2010 Aug;9(8):596. doi: 10.1038/nrd3234. Nat Rev Drug Discov. 2010. PMID: 20671763 No abstract available.
-
Spinal cord injury: The two faces of spontaneous serotonin signalling.Nat Rev Neurosci. 2010 Aug;11(8):535. doi: 10.1038/nrn2888. Nat Rev Neurosci. 2010. PMID: 20672426 No abstract available.
References
-
- Carlsson A, Magnusson T, Rosengren E. 5-hydroxytryptamine of the spinal cord normally and after transection. Experientia. 1963;19:359. - PubMed
-
- Jacobs BL, Martin-Cora FJ, Fornal CA. Activity of medullary serotonergic neurons in freely moving animals. Brain Res. Brain Res. Rev. 2002;40:45–52. - PubMed
-
- Jordan LM, Liu J, Hedlund PB, Akay T, Pearson KG. Descending command systems for the initiation of locomotion in mammals. Brain Res. Rev. 2008;57:183–191. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
