

Activating WASP mutations associated with X-linked neutropenia result in enhanced actin polymerization, altered cytoskeletal responses, and genomic instability in lymphocytes
- PMID: 20513746
- PMCID: PMC2882832
- DOI: 10.1084/jem.20091245
Activating WASP mutations associated with X-linked neutropenia result in enhanced actin polymerization, altered cytoskeletal responses, and genomic instability in lymphocytes
Abstract
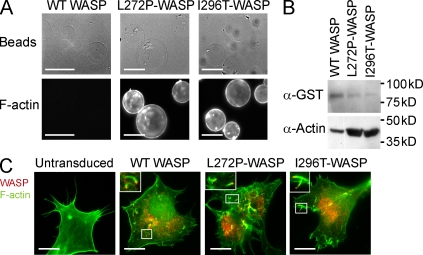
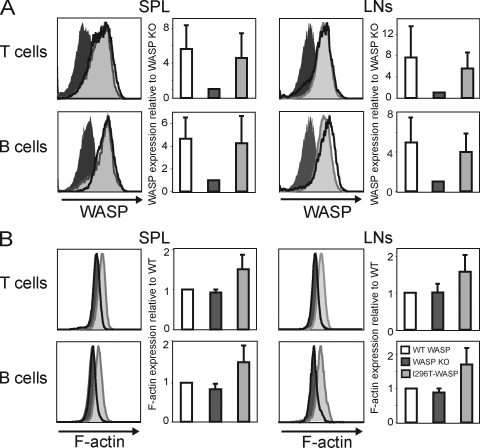
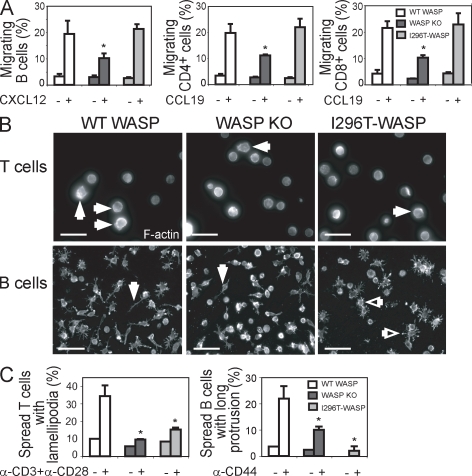
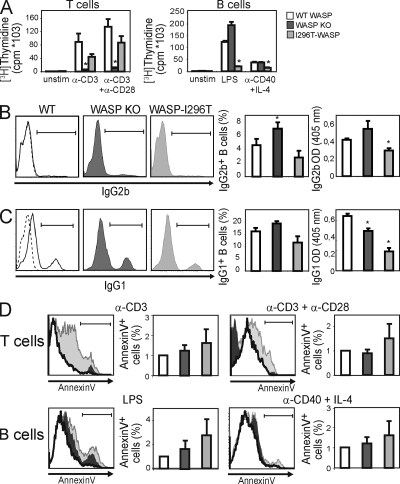
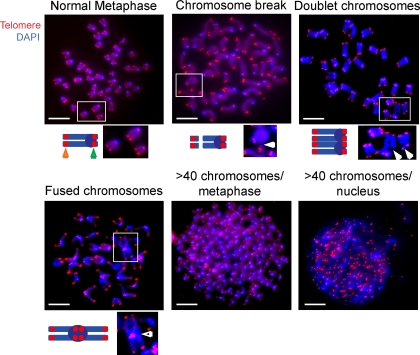
X-linked neutropenia (XLN) is caused by activating mutations in the Wiskott-Aldrich syndrome protein (WASP) that result in aberrant autoinhibition. Although patients with XLN appear to have only defects in myeloid lineages, we hypothesized that activating mutations of WASP are likely to affect the immune system more broadly. We generated mouse models to assess the role of activating WASP mutations associated with XLN (XLN-WASP) in lymphocytes. XLN-WASP is expressed stably in B and T cells and induces a marked increase in polymerized actin. XLN-WASP-expressing B and T cells migrate toward chemokines but fail to adhere normally. In marked contrast to WASP-deficient cells, XLN-WASP-expressing T cells proliferate normally in response to cell-surface receptor activation. However, XLN-WASP-expressing B cells fail to proliferate and secrete lower amounts of antibodies. Moreover, XLN-WASP expression in lymphocytes results in modestly increased apoptosis associated with increased genomic instability. These data indicate that there are unique requirements for the presence and activation status of WASP in B and T cells and that WASP-activating mutations interfere with lymphocyte cell survival and genomic stability.
Figures
References
-
- Ancliff P.J., Blundell M.P., Cory G.O., Calle Y., Worth A., Kempski H., Burns S., Jones G.E., Sinclair J., Kinnon C., et al. 2006. Two novel activating mutations in the Wiskott-Aldrich syndrome protein result in congenital neutropenia. Blood. 108:2182–2189 10.1182/blood-2006-01-010249 - DOI - PubMed
-
- Beel K., Cotter M.M., Blatny J., Bond J., Lucas G., Green F., Vanduppen V., Leung D.W., Rooney S., Smith O.P., et al. 2009. A large kindred with X-linked neutropenia with an I294T mutation of the Wiskott-Aldrich syndrome gene. Br. J. Haematol. 144:120–126 10.1111/j.1365-2141.2008.07416.x - DOI - PMC - PubMed
