

The tumor suppressor p53: from structures to drug discovery
- PMID: 20516128
- PMCID: PMC2869527
- DOI: 10.1101/cshperspect.a000919
The tumor suppressor p53: from structures to drug discovery
Abstract
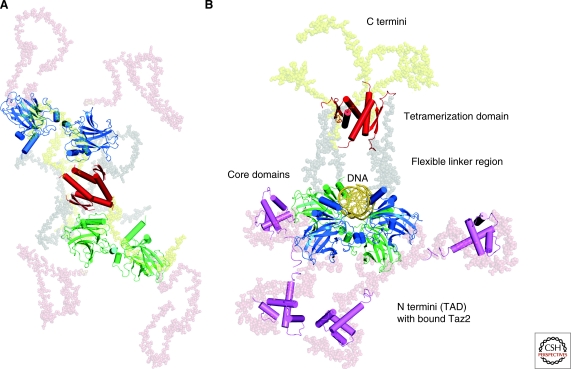
Even 30 years after its discovery, the tumor suppressor protein p53 is still somewhat of an enigma. p53's intimate and multifaceted role in the cell cycle is mirrored in its equally complex structural biology that is being unraveled only slowly. Here, we discuss key structural aspects of p53 function and its inactivation by oncogenic mutations. Concerted action of folded and intrinsically disordered domains of the highly dynamic p53 protein provides binding promiscuity and specificity, allowing p53 to process a myriad of cellular signals to maintain the integrity of the human genome. Importantly, progress in elucidating the structural biology of p53 and its partner proteins has opened various avenues for structure-guided rescue of p53 function in tumors. These emerging anticancer strategies include targeting mutant-specific lesions on the surface of destabilized cancer mutants with small molecules and selective inhibition of p53's degradative pathways.
Figures





References
-
- Achatz MI, Olivier M, Le Calvez F, Martel-Planche G, Lopes A, Rossi BM, Ashton-Prolla P, Giugliani R, Palmero EI, Vargas FR, et al.2007. The TP53 mutation, R337H, is associated with Li-Fraumeni and Li-Fraumeni-like syndromes in Brazilian families. Cancer Lett 245:96–102 - PubMed
-
- Ang HC, Joerger AC, Mayer S, Fersht AR 2006. Effects of common cancer mutations on stability and DNA binding of full-length p53 compared with isolated core domains. J Biol Chem 281:21934–21941 - PubMed
-
- Avalos JL, Celic I, Muhammad S, Cosgrove MS, Boeke JD, Wolberger C 2002. Structure of a Sir2 enzyme bound to an acetylated p53 peptide. Mol Cell 10:523–535 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous