

Review
doi: 10.1101/cshperspect.a001966.
Epub 2009 Dec 23.
Use of forward genetics to discover novel regulators of NF-kappaB
Affiliations
- PMID: 20516132
- PMCID: PMC2869522
- DOI: 10.1101/cshperspect.a001966
Item in Clipboard
Review
Use of forward genetics to discover novel regulators of NF-kappaB
Cold Spring Harb Perspect Biol.
2010 Jun.
Abstract
Forward and reverse genetic experiments have both played important roles in revealing critical aspects of mammalian signal transduction pathways in cell culture experiments. Only recently have we begun to comprehend the depth, breadth, and complexity of these pathways and of their interrelationships. Here, we summarize successful examples in which different forward genetic approaches have led to novel discoveries in NF-kappaB signaling. We believe that forward genetics will continue to play an irreplaceable role in advancing our understanding of the complexities of the pathways that regulate the functions of this key transcription factor.
Figures
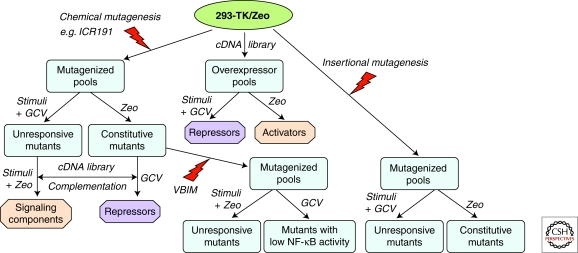
General scheme for forward genetics analysis of NF-κB-dependent signaling pathway, using lethal selection to identify regulative proteins. 293-TK/Zeo cells, carrying both TK and Zeo selectable markers, can be used in conjunction with chemical mutagenesis (left) or insertional mutagenesis (right) to obtain mutants unresponsive to a stimulus or constitutive mutants, and then to identify either stimulus-specific signaling components or general activators or repressors. A cDNA library (middle) can also be overexpressed to screen directly for activators or repressors. Additionally, constitutive mutants can be further mutated by insertional mutagenesis using VBIM (bottom) to screen for mutants with low NF-κB to identify activators or repressors.
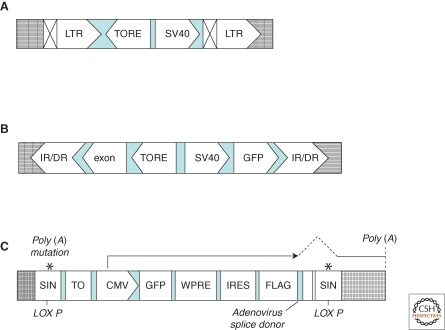
Structures of different promoter insertion vectors. (A) Retroviral vectors. Retroviral vectors optimized for insertional mutagenesis were generated by using the MMLV (Moloney murine leukemia virus) backbone of pBabe vectors. The recognition sequence for Cre recombinase was placed inside the 3′ LTR so that, on reverse transcription, this motif would be copied into the 5′-LTR (Kandel et al. 2005). Abbreviations: TORE, tetracycline-regulated promoter; SV40, promoter and enhancer from the SV40 virus; LTR, long terminal repeat. Symbols: ⊠, the LTR modification, including a LoxP site for cre-mediated recombination, leading to deletion of the promoter; grid lines, host genomic DNA. (B) Sleeping Beauty transposon-based vectors. This series of mutagenesis vectors contain a coding region for GFP controlled by TORE, which is placed in the vicinity of a divergent SV40 promoter, followed by a mini-exon, which ends with an unpaired adenoviral splice site (Kandel et al. 2005). These vectors can be cotransfected with the Sleeping Beauty transposase to randomly generate mutations. Because the promoter is tet-regulated, mutant phenotypes caused by the inserted promoter can be reverted on promoter shutdown on treatment with the tetracycline analog doxycycline (Dasgupta et al. 2008). Abbreviations: IR/DR, terminal repeat from Sleeping Beauty; TORE, modified tetracycline-regulated promoter; GFP, coding region of enhanced GFP. Symbol: grid lines, host genomic DNA. (C) VBIM vectors. The VBIM vectors use a lentiviral backbone with polyadenylation mutations in both 5′ and 3′ LTRs and a lox P site in the 3′ LTR. This design allows excision of all but 238 bp of inert proviral DNA, lacking both promoter activity and polyadenylation signals, following cleavage by Cre, a critical feature for complete and consistent phenotypic reversion. The polyadenylation mutations also permit the mutagenic promoter to be placed in the same direction as transcription from the 5′ LTR during virus packaging, resulting in high virus titers that are comparable to those obtained with standard lentiviral vectors, eliminating promoter conflicts that occur with alternative designs. Furthermore, eliminating promoter interference also permits the use of a strong full-length CMV promoter rather than a minimal tetracycline-regulated promoter, which requires the tet activator protein tTA to be present in the target cells (Kandel et al. 2005). Thus, primary and even differentiated or senescent cells can be mutated without prior manipulation to express tTA or an ecotropic receptor, as previously required (Kandel et al. 2005). Besides these, a tetracycline-binding element upstream of the full-length CMV promoter is introduced to allow tetracycline-regulated control of the mutagenic promoter after the mutant has been created, by using a TR-KRAB fusion protein. Detailed description of this series of vectors has been reported recently (Lu et al. 2009a). Abbreviations: CMV, cytomegalovirus promoter; GFP, green fluorescent protein; IRES, internal ribosome entry sequence; LoxP, site for cre-mediated recombination; SIN, self-inactivating LTR; TO, tetracycline operon; WPRE, woodchuck hepatitis virus post-transcriptional regulatory element. Symbol: grid lines, host genomic DNA.
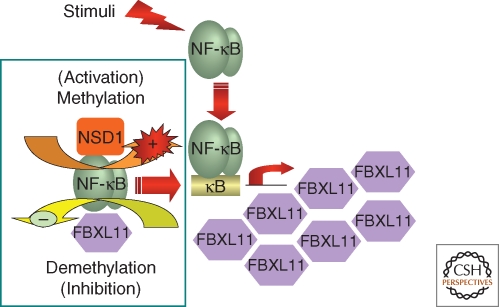
Regulation of NF-κB by lysine methylation. NF-κB is regulated through reversible methylation of p65, catalyzed by the NSD1–FBXL11 enzyme pair. The FBXL11 gene is activated by NF-κB, forming a negative-feedback loop that down-regulates NF-κB.
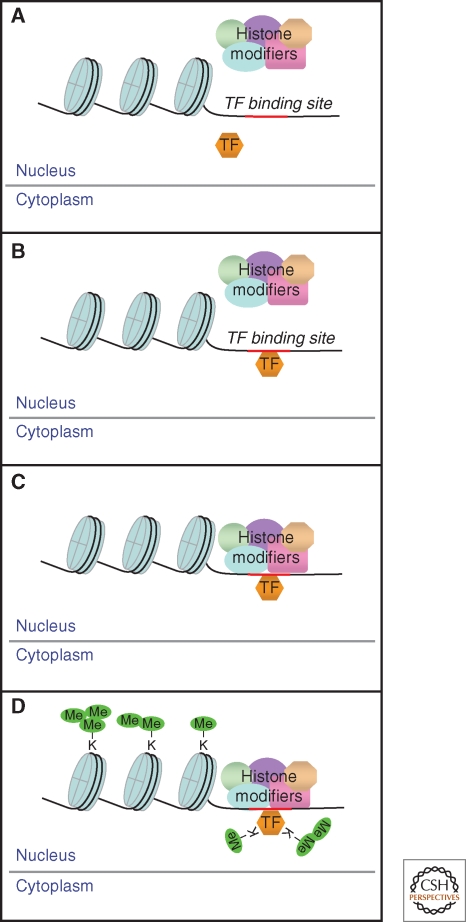
Model for transcription factor methylation by histone-modifying enzymes. (A) The activated transcription factor (TF) enters the nucleus and (B) binds to a promoter. (C) The histone-modifying complex is recruited. (D) This complex then methylates both the histones and the promoter-bound TF. Multiple sites are mono-, di-, and tri-methylated, profoundly affecting the functions of both the histones and the TF.
Similar articles
-
Signaling to NF-kappaB.Genes Dev. 2004 Sep 15;18(18):2195-224. doi: 10.1101/gad.1228704. Genes Dev. 2004. PMID: 15371334 Review.
-
Computational modeling with forward and reverse engineering links signaling network and genomic regulatory responses: NF-kappaB signaling-induced gene expression responses in inflammation.BMC Bioinformatics. 2010 Jun 8;11:308. doi: 10.1186/1471-2105-11-308. BMC Bioinformatics. 2010. PMID: 20529327 Free PMC article.
-
The nuclear factor NF-kappaB pathway in inflammation.Cold Spring Harb Perspect Biol. 2009 Dec;1(6):a001651. doi: 10.1101/cshperspect.a001651. Epub 2009 Oct 7. Cold Spring Harb Perspect Biol. 2009. PMID: 20457564 Free PMC article. Review.
-
The NF-κB Family of Transcription Factors and Its Role in Thyroid Physiology.Vitam Horm. 2018;106:195-210. doi: 10.1016/bs.vh.2017.05.003. Epub 2017 Sep 4. Vitam Horm. 2018. PMID: 29407436
-
Transmembrane TNF-alpha mediates "forward" and "reverse" signaling, inducing cell death or survival via the NF-kappaB pathway in Raji Burkitt lymphoma cells.J Leukoc Biol. 2008 Sep;84(3):789-97. doi: 10.1189/jlb.0208078. Epub 2008 Jun 11. J Leukoc Biol. 2008. PMID: 18550789 Free PMC article.
Cited by
-
Identification of an N-terminal truncation of the NF-κB p65 subunit that specifically modulates ribosomal protein S3-dependent NF-κB gene expression.J Biol Chem. 2012 Dec 14;287(51):43019-29. doi: 10.1074/jbc.M112.388694. Epub 2012 Oct 31. J Biol Chem. 2012. PMID: 23115242 Free PMC article.
-
NF-κB: Regulation by Methylation.Cancer Res. 2015 Sep 15;75(18):3692-5. doi: 10.1158/0008-5472.CAN-15-1022. Epub 2015 Sep 3. Cancer Res. 2015. PMID: 26337909 Free PMC article. Review.
-
Other ways to skin a cat: activating SREBP without Scap.Fly (Austin). 2011 Jan-Mar;5(1):3-6. doi: 10.4161/fly.5.1.13475. Epub 2011 Jan 1. Fly (Austin). 2011. PMID: 20935466 Free PMC article.
-
Discovery and characterization of anti-cancer peptides from a random peptide library.PLoS One. 2024 Feb 13;19(2):e0293072. doi: 10.1371/journal.pone.0293072. eCollection 2024. PLoS One. 2024. PMID: 38349913 Free PMC article.
-
Forward genetic screening for regulators involved in cholesterol synthesis using validation-based insertional mutagenesis.PLoS One. 2014 Nov 26;9(11):e112632. doi: 10.1371/journal.pone.0112632. eCollection 2014. PLoS One. 2014. PMID: 25426949 Free PMC article.
References
-
- Banks DJ, Bradley KA 2007. SILENCE: A new forward genetic technology. Nat Methods 4:51–53 - PubMed
-
- Berns K, Hijmans EM, Mullenders J, Brummelkamp TR, Velds A, Heimerikx M, Kerkhoven RM, Madiredjo M, Nijkamp W, Weigelt B, et al.2004. A large-scale RNAi screen in human cells identifies new components of the p53 pathway. Nature 428:431–437 - PubMed
-
- Cao Z, Henzel WJ, Gao X 1996. IRAK: A kinase associated with the interleukin-1 receptor. Science 271:1128–1131 - PubMed
-
- Collier LS, Carlson CM, Ravimohan S, Dupuy AJ, Largaespada DA 2005. Cancer gene discovery in solid tumours using transposon-based somatic mutagenesis in the mouse. Nature 436:272–276 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources