

Distinct roles for PTEN in prevention of T cell lymphoma and autoimmunity in mice
- PMID: 20516645
- PMCID: PMC2898609
- DOI: 10.1172/JCI42382
Distinct roles for PTEN in prevention of T cell lymphoma and autoimmunity in mice
Abstract
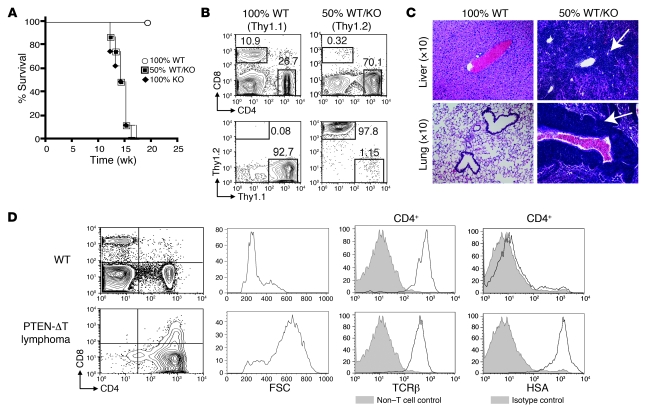
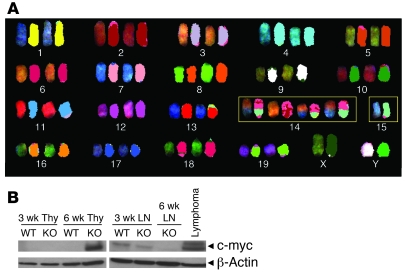
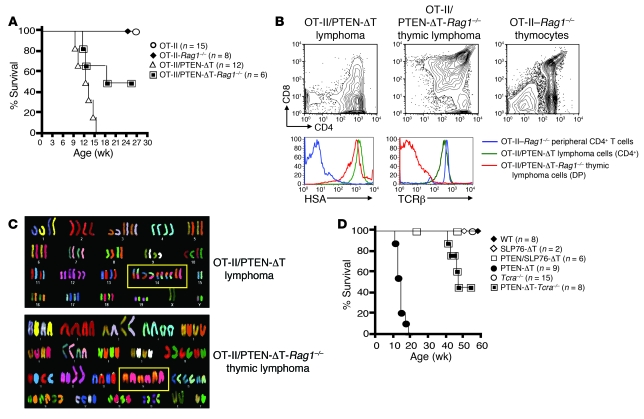
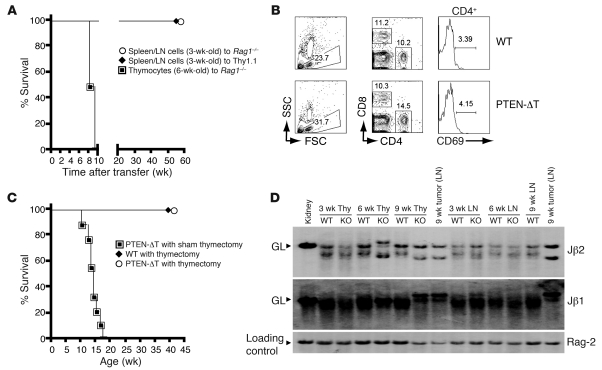
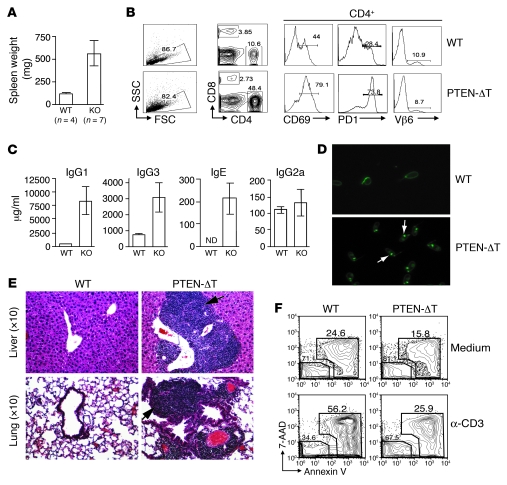
Mutations in the tumor-suppressor gene phosphatase and tensin homolog deleted on chromosome 10 (Pten) are associated with multiple cancers in humans, including T cell malignancies. Targeted deletion of Pten in T cells induces both a disseminated "mature phenotype" lymphoma and a lymphoproliferative autoimmune syndrome in mice. Here, we have shown that these two diseases are separable and mediated by T lineage cells of distinct developmental stages. Loss of PTEN was found to be a powerful driver of lymphomagenesis within the thymus characterized by overexpression of the c-myc oncogene. In an otherwise normal thymic environment, PTEN-deficient T cell lymphomas invariably harbored RAG-dependent reciprocal t(14:15) chromosomal translocations involving the T cell receptor alpha/delta locus and c-myc, and their survival and growth was TCR dependent, but Notch independent. However, lymphomas occurred even if TCR recombination was prevented, although these lymphomas were less mature, arose later in life, and, importantly, were dependent upon Notch pathways to upregulate c-myc expression. In contrast, using the complementary methods of early thymectomy and adoptive transfers, we found that PTEN-deficient mature T cells were unable to undergo malignant transformation but were sufficient for the development of autoimmunity. These data suggest multiple and distinct regulatory roles for PTEN in the molecular pathogenesis of lymphoma and autoimmunity.
Figures

References
-
- Li J, et al. PTEN, a putative protein tyrosine phosphatase gene mutated in human brain, breast, and prostate cancer. Science. 1997;275(5308):1943–1947. - PubMed
-
- Maehama T, Dixon JE. The tumor suppressor, PTEN/MMAC1, dephosphorylates the lipid second messenger, phosphatidylinositol 3,4,5-trisphosphate. J Biol Chem. 1998;273(22):13375–13378. - PubMed
-
- Engelman JA, Luo J, Cantley LC. The evolution of phosphatidylinositol 3-kinases as regulators of growth and metabolism. Nat Rev Genet. 2006;7(8):606–619. - PubMed
-
- Teng DH, et al. MMAC1/PTEN mutations in primary tumor specimens and tumor cell lines. Cancer Res. 1997;57(23):5221–5225. - PubMed
-
- Liaw D, et al. Germline mutations of the PTEN gene in Cowden disease, an inherited breast and thyroid cancer syndrome. Nat Genet. 1997;16(1):64–67. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
