

Neurokinin B acts via the neurokinin-3 receptor in the retrochiasmatic area to stimulate luteinizing hormone secretion in sheep
- PMID: 20519368
- PMCID: PMC2940514
- DOI: 10.1210/en.2010-0174
Neurokinin B acts via the neurokinin-3 receptor in the retrochiasmatic area to stimulate luteinizing hormone secretion in sheep
Abstract
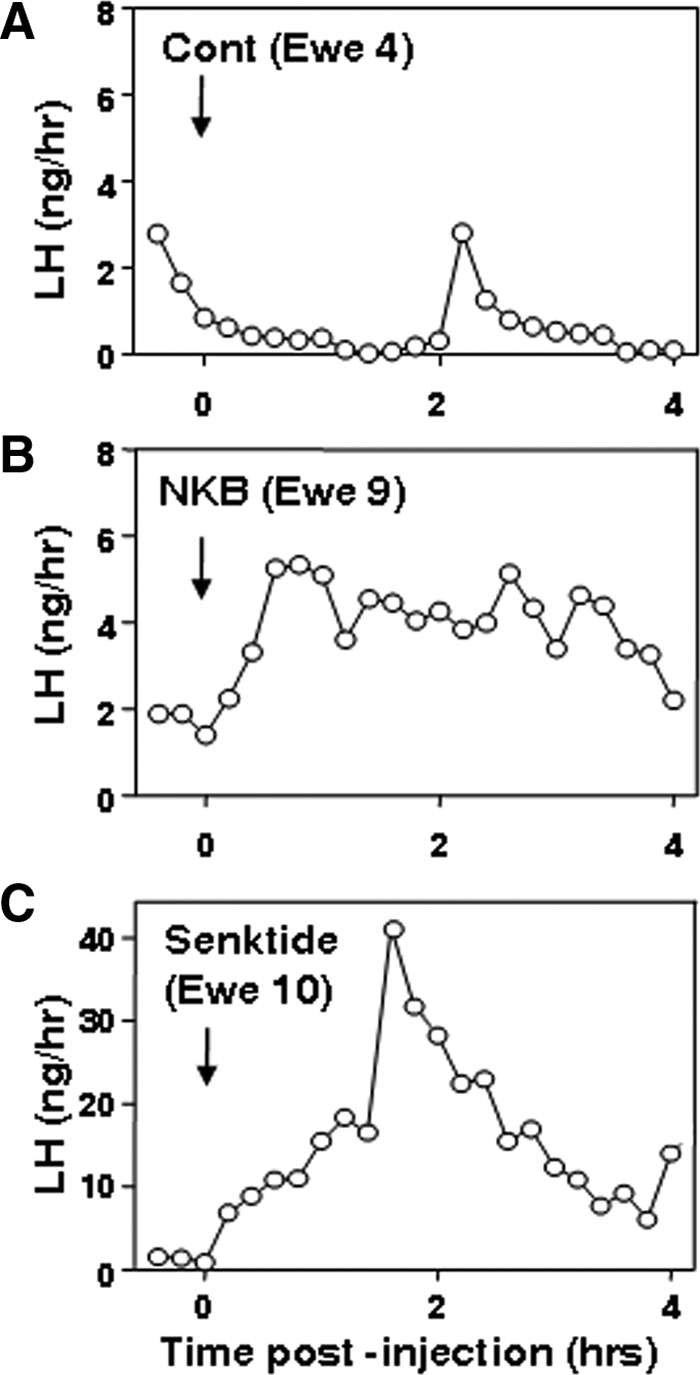
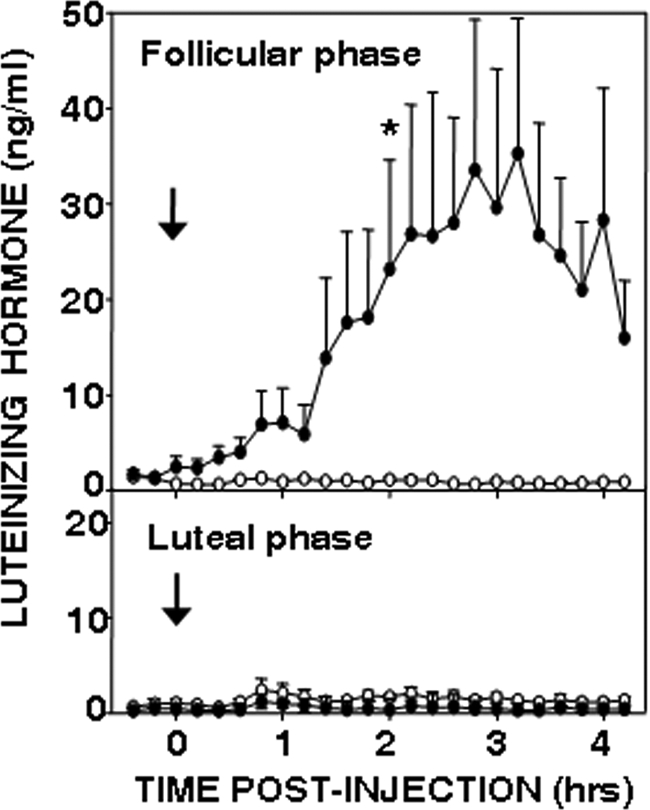
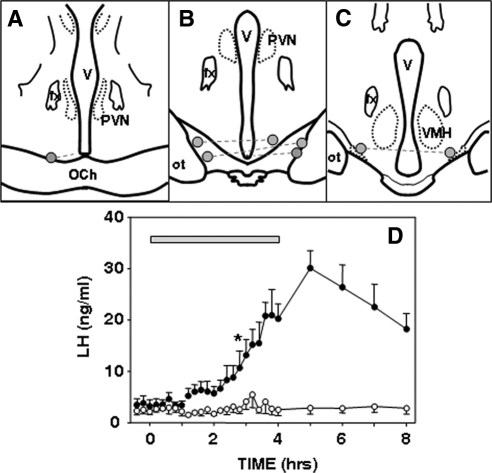
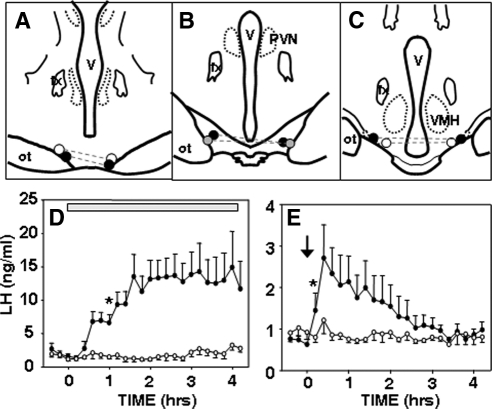
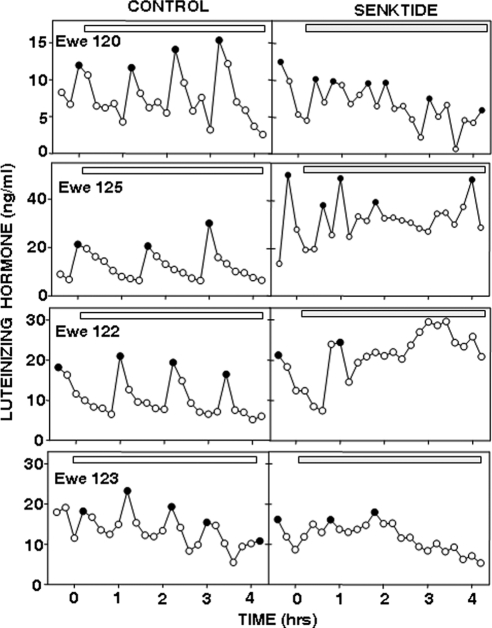
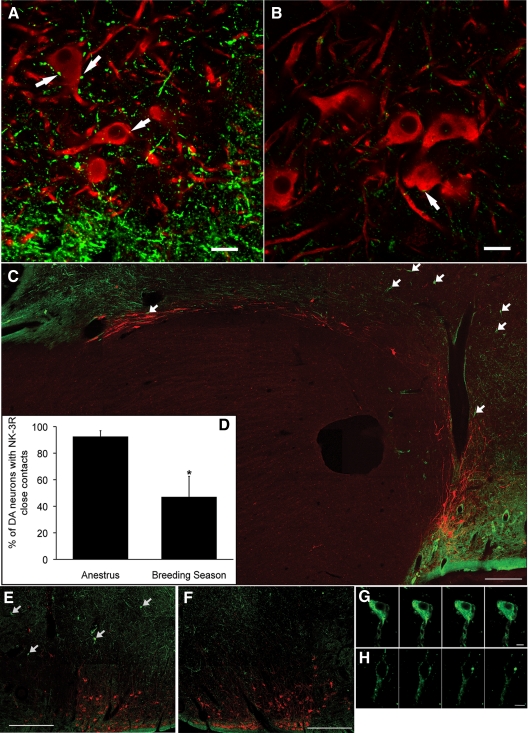
Recent data have demonstrated that mutations in the receptor for neurokinin B (NKB), the NK-3 receptor (NK3R), produce hypogonadotropic hypogonadism in humans. These data, together with reports that NKB expression increases after ovariectomy and in postmenopausal women, have led to the hypothesis that this tachykinin is an important stimulator of GnRH secretion. However, the NK3R agonist, senktide, inhibited LH secretion in rats and mice. In this study, we report that senktide stimulates LH secretion in ewes. A dramatic increase in LH concentrations to levels close to those observed during the preovulatory LH surge was observed after injection of 1 nmol senktide into the third ventricle during the follicular, but not in the luteal, phase. Similar increases in LH secretion occurred after insertion of microimplants containing this agonist into the retrochiasmatic area (RCh) in anestrous or follicular phase ewes. A low-dose microinjection (3 pmol) of senktide into the RCh produced a smaller but significant increase in LH concentrations in anestrous ewes. Moreover, NK3R immunoreactivity was clearly evident in the RCh, although it was not found in A15 dopaminergic cell bodies in this region. These data provide evidence that NKB stimulates LH (and presumably GnRH) secretion in ewes and point to the RCh as one important site of action. Based on these data, and the effects of NK3R mutations in humans, we hypothesize that NKB plays an important stimulatory role in the control of GnRH and LH secretion in nonrodent species.
Figures
References
-
- Seminara SB, Messager S, Chatzidaki EE, Thresher RR, Acierno Jr JS, Shagoury JK, Bo-Abbas Y, Kuohung W, Schwinof KM, Hendrick AG, Zahn D, Dixon J, Kaiser UB, Slaugenhaupt SA, Gusella JF, O'Rahilly S, Carlton MB, Crowley Jr WF, Aparicio SA, Colledge WH 2003 The GPR54 gene as a regulator of puberty. N Engl J Med 349:1614–1627 - PubMed
-
- Smith JT 2009 Sex steroid control of hypothalamic Kiss 1 expression in sheep and rodents: comparative aspects. Peptides 30:94–102 - PubMed
