

Vaccination with murid herpesvirus-4 glycoprotein B reduces viral lytic replication but does not induce detectable virion neutralization
- PMID: 20519454
- PMCID: PMC3052599
- DOI: 10.1099/vir.0.023085-0
Vaccination with murid herpesvirus-4 glycoprotein B reduces viral lytic replication but does not induce detectable virion neutralization
Abstract
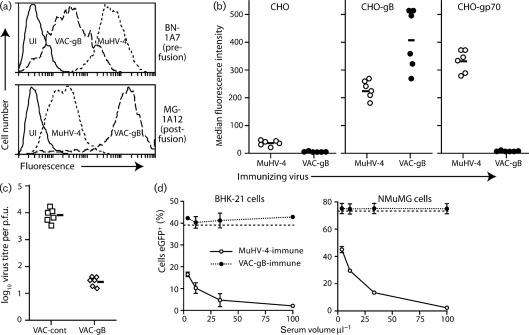
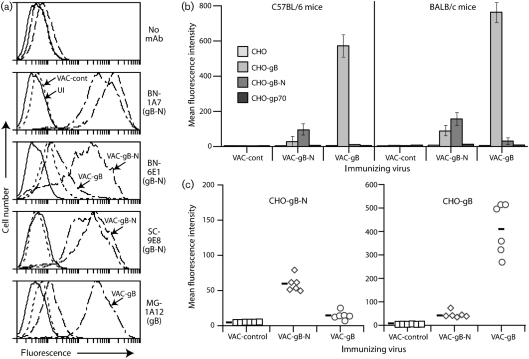
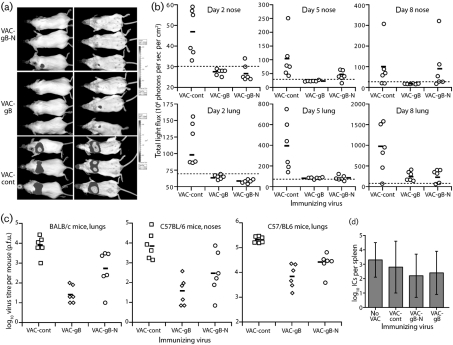
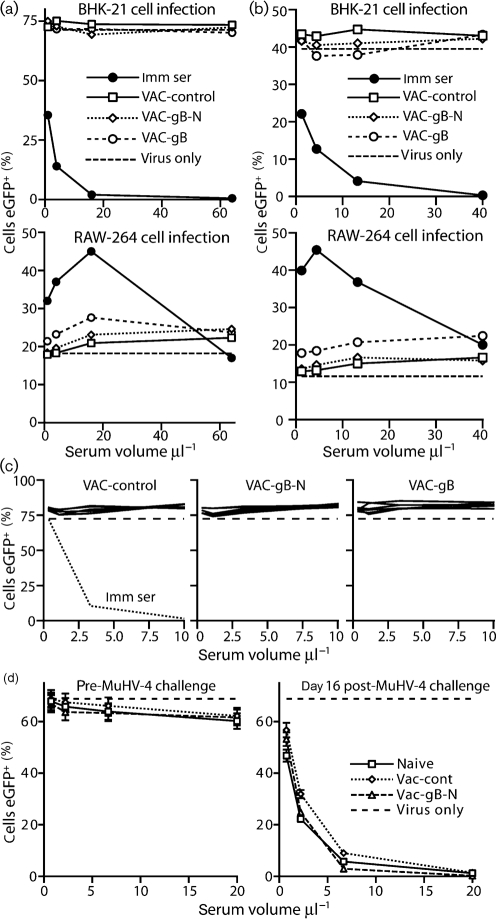
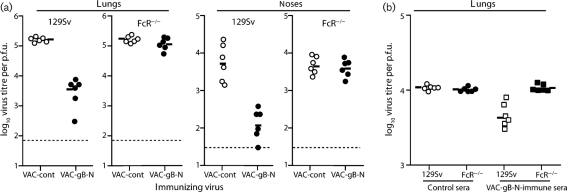
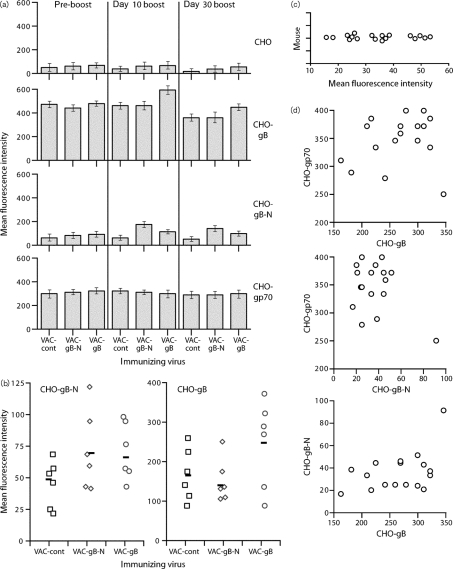
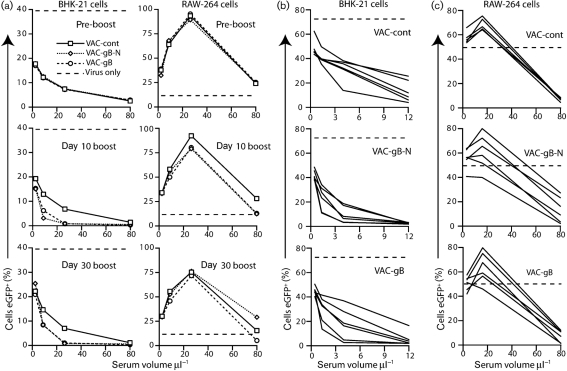
Herpesviruses characteristically disseminate from immune hosts. Therefore in the context of natural infection, antibody neutralizes them poorly. Murid herpesvirus-4 (MuHV-4) provides a tractable model with which to understand gammaherpesvirus neutralization. MuHV-4 virions blocked for cell binding by immune sera remain infectious for IgG-Fc receptor(+) myeloid cells, so broadly neutralizing antibodies must target the virion fusion complex - glycoprotein B (gB) or gH/gL. While gB-specific neutralizing antibodies are rare, its domains I+II (gB-N) contain at least one potent neutralization epitope. Here, we tested whether immunization with recombinant gB presenting this epitope could induce neutralizing antibodies in naive mice and protect them against MuHV-4 challenge. Immunizing with the full-length gB extracellular domain induced a strong gB-specific antibody response and reduced MuHV-4 lytic replication but did not induce detectable neutralization. gB-N alone, which more selectively displayed pre-fusion epitopes including neutralization epitopes, also failed to induce neutralizing responses, and while viral lytic replication was again reduced this depended completely on IgG Fc receptors. gB and gB-N also boosted neutralizing responses in only a minority of carrier mice. Therefore, it appears that neutralizing epitopes on gB are intrinsically difficult for the immune response to target.
Figures
Similar articles
-
Glycoprotein L sets the neutralization profile of murid herpesvirus 4.J Gen Virol. 2009 May;90(Pt 5):1202-1214. doi: 10.1099/vir.0.008755-0. Epub 2009 Mar 4. J Gen Virol. 2009. PMID: 19264603 Free PMC article.
-
A mechanistic basis for potent, glycoprotein B-directed gammaherpesvirus neutralization.J Gen Virol. 2011 Sep;92(Pt 9):2020-2033. doi: 10.1099/vir.0.032177-0. Epub 2011 May 18. J Gen Virol. 2011. PMID: 21593277 Free PMC article.
-
Virion endocytosis is a major target for murid herpesvirus-4 neutralization.J Gen Virol. 2012 Jun;93(Pt 6):1316-1327. doi: 10.1099/vir.0.040790-0. Epub 2012 Feb 29. J Gen Virol. 2012. PMID: 22377583 Free PMC article.
-
Antibody limits in vivo murid herpesvirus-4 replication by IgG Fc receptor-dependent functions.J Gen Virol. 2009 Nov;90(Pt 11):2592-2603. doi: 10.1099/vir.0.014266-0. Epub 2009 Jul 22. J Gen Virol. 2009. PMID: 19625459 Free PMC article.
-
Immune control of mammalian gamma-herpesviruses: lessons from murid herpesvirus-4.J Gen Virol. 2009 Oct;90(Pt 10):2317-2330. doi: 10.1099/vir.0.013300-0. Epub 2009 Jul 15. J Gen Virol. 2009. PMID: 19605591 Review.
Cited by
-
Type I Interferons Direct Gammaherpesvirus Host Colonization.PLoS Pathog. 2016 May 25;12(5):e1005654. doi: 10.1371/journal.ppat.1005654. eCollection 2016 May. PLoS Pathog. 2016. PMID: 27223694 Free PMC article.
-
ORF6 and ORF61 Expressing MVA Vaccines Impair Early but Not Late Latency in Murine Gammaherpesvirus MHV-68 Infection.Front Immunol. 2019 Dec 18;10:2984. doi: 10.3389/fimmu.2019.02984. eCollection 2019. Front Immunol. 2019. PMID: 31921215 Free PMC article.
-
CD8+ T cell evasion mandates CD4+ T cell control of chronic gamma-herpesvirus infection.PLoS Pathog. 2017 Apr 10;13(4):e1006311. doi: 10.1371/journal.ppat.1006311. eCollection 2017 Apr. PLoS Pathog. 2017. PMID: 28394921 Free PMC article.
-
Herpes simplex virus 1 targets the murine olfactory neuroepithelium for host entry.J Virol. 2013 Oct;87(19):10477-88. doi: 10.1128/JVI.01748-13. Epub 2013 Jul 31. J Virol. 2013. PMID: 23903843 Free PMC article.
-
Gammaherpesvirus Colonization of the Spleen Requires Lytic Replication in B Cells.J Virol. 2018 Mar 14;92(7):e02199-17. doi: 10.1128/JVI.02199-17. Print 2018 Apr 1. J Virol. 2018. PMID: 29343572 Free PMC article.
References
-
- Akula, S. M., Pramod, N. P., Wang, F. Z. & Chandran, B. (2001). Human herpesvirus 8 envelope-associated glycoprotein B interacts with heparan sulfate-like moieties. Virology 284, 235–249. - PubMed
-
- Cranage, M. P., Kouzarides, T., Bankier, A. T., Satchwell, S., Weston, K., Tomlinson, P., Barrell, B., Hart, H., Bell, S. E. & other authors (1986). Identification of the human cytomegalovirus glycoprotein B gene and induction of neutralizing antibodies via its expression in recombinant vaccinia virus. EMBO J 5, 3057–3063. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources