

Roles of platelet STIM1 and Orai1 in glycoprotein VI- and thrombin-dependent procoagulant activity and thrombus formation
- PMID: 20519511
- PMCID: PMC2911319
- DOI: 10.1074/jbc.M110.108696
Roles of platelet STIM1 and Orai1 in glycoprotein VI- and thrombin-dependent procoagulant activity and thrombus formation
Abstract
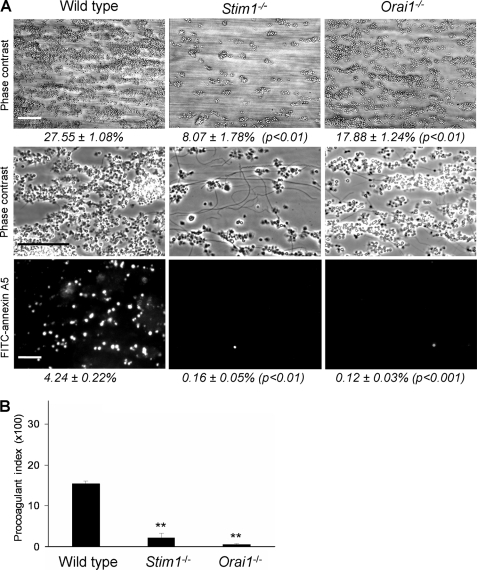
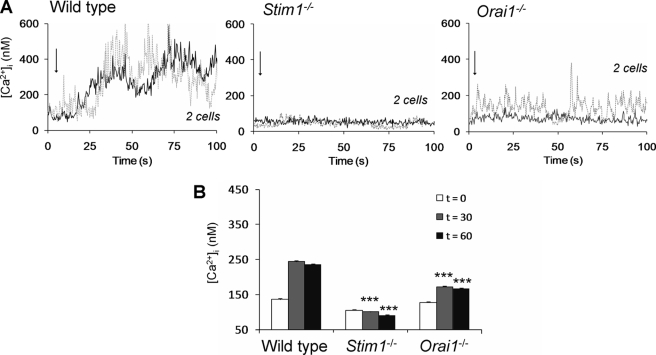
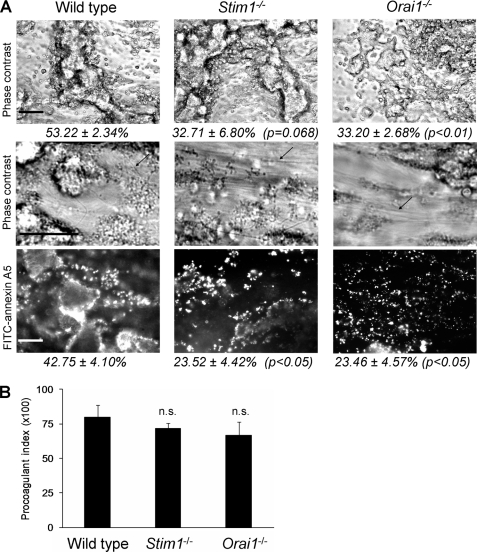
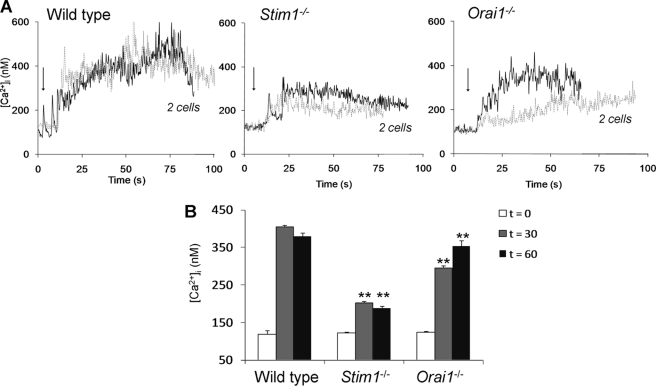
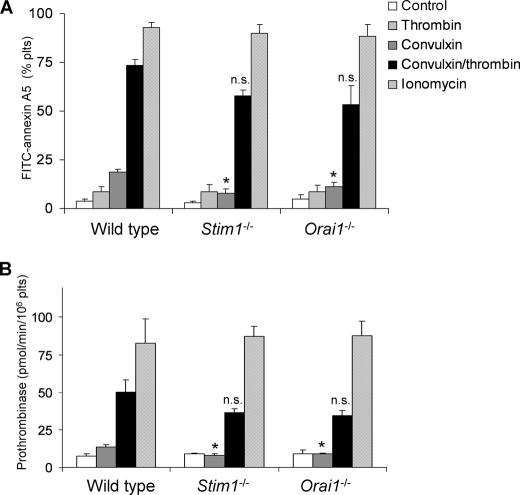
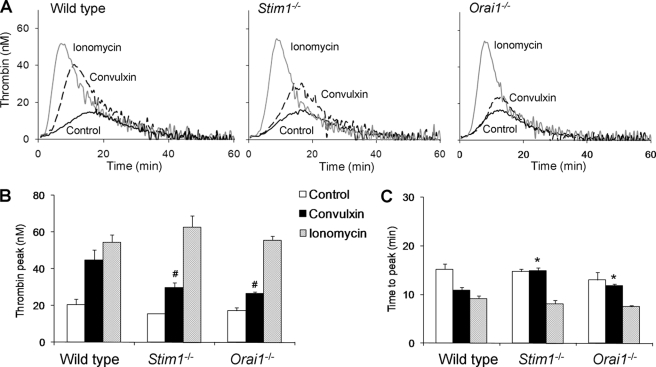
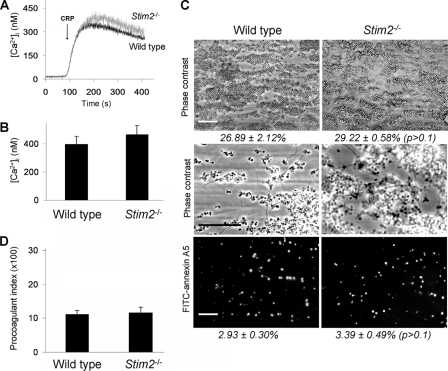
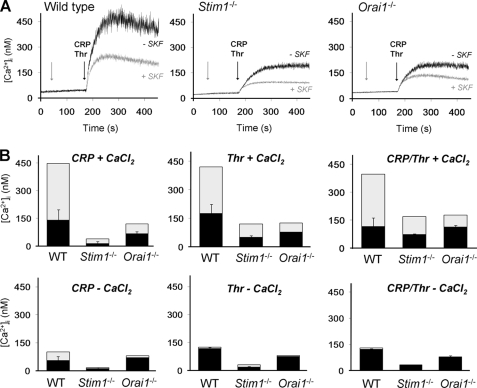
In platelets, STIM1 has been recognized as the key regulatory protein in store-operated Ca(2+) entry (SOCE) with Orai1 as principal Ca(2+) entry channel. Both proteins contribute to collagen-dependent arterial thrombosis in mice in vivo. It is unclear whether STIM2 is involved. A key platelet response relying on Ca(2+) entry is the surface exposure of phosphatidylserine (PS), which accomplishes platelet procoagulant activity. We studied this response in mouse platelets deficient in STIM1, STIM2, or Orai1. Upon high shear flow of blood over collagen, Stim1(-/-) and Orai1(-/-) platelets had greatly impaired glycoprotein (GP) VI-dependent Ca(2+) signals, and they were deficient in PS exposure and thrombus formation. In contrast, Stim2(-/-) platelets reacted normally. Upon blood flow in the presence of thrombin generation and coagulation, Ca(2+) signals of Stim1(-/-) and Orai1(-/-) platelets were partly reduced, whereas the PS exposure and formation of fibrin-rich thrombi were normalized. Washed Stim1(-/-) and Orai1(-/-) platelets were deficient in GPVI-induced PS exposure and prothrombinase activity, but not when thrombin was present as co-agonist. Markedly, SKF96365, a blocker of (receptor-operated) Ca(2+) entry, inhibited Ca(2+) and procoagulant responses even in Stim1(-/-) and Orai1(-/-) platelets. These data show for the first time that: (i) STIM1 and Orai1 jointly contribute to GPVI-induced SOCE, procoagulant activity, and thrombus formation; (ii) a compensating Ca(2+) entry pathway is effective in the additional presence of thrombin; (iii) platelets contain two mechanisms of Ca(2+) entry and PS exposure, only one relying on STIM1-Orai1 interaction.
Figures
References
-
- Heemskerk J. W., Bevers E. M., Lindhout T. (2002) Thromb. Haemost. 88, 186–193 - PubMed
-
- Heemskerk J. W., Feijge M. A., Henneman L., Rosing J., Hemker H. C. (1997) Eur. J. Biochem. 249, 547–555 - PubMed
-
- Zwaal R. F., Schroit A. J. (1997) Blood 89, 1121–1132 - PubMed
-
- Heemskerk J. W., Kuijpers M. J., Munnix I. C., Siljander P. R. (2005) Trends Cardiovasc. Med. 15, 86–92 - PubMed
-
- Munnix I. C., Strehl A., Kuijpers M. J., Auger J. M., van der Meijden P. E., van Zandvoort M. A., oude Egbrink M., Nieswandt B., Heemskerk J. W. (2005) Arterioscler. Thromb. Vasc. Biol. 25, 2673–2678 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
