

Review
doi: 10.1038/nrm2917.
Epub 2010 Jun 3.
All things must pass: contrasts and commonalities in eukaryotic and bacterial mRNA decay
Affiliations
- PMID: 20520623
- PMCID: PMC3145457
- DOI: 10.1038/nrm2917
Item in Clipboard
Review
All things must pass: contrasts and commonalities in eukaryotic and bacterial mRNA decay
Nat Rev Mol Cell Biol.
2010 Jul.
Abstract
Despite its universal importance for controlling gene expression, mRNA degradation was initially thought to occur by disparate mechanisms in eukaryotes and bacteria. This conclusion was based on differences in the structures used by these organisms to protect mRNA termini and in the RNases and modifying enzymes originally implicated in mRNA decay. Subsequent discoveries have identified several striking parallels between the cellular factors and molecular events that govern mRNA degradation in these two kingdoms of life. Nevertheless, some key distinctions remain, the most fundamental of which may be related to the different mechanisms by which eukaryotes and bacteria control translation initiation.
Figures
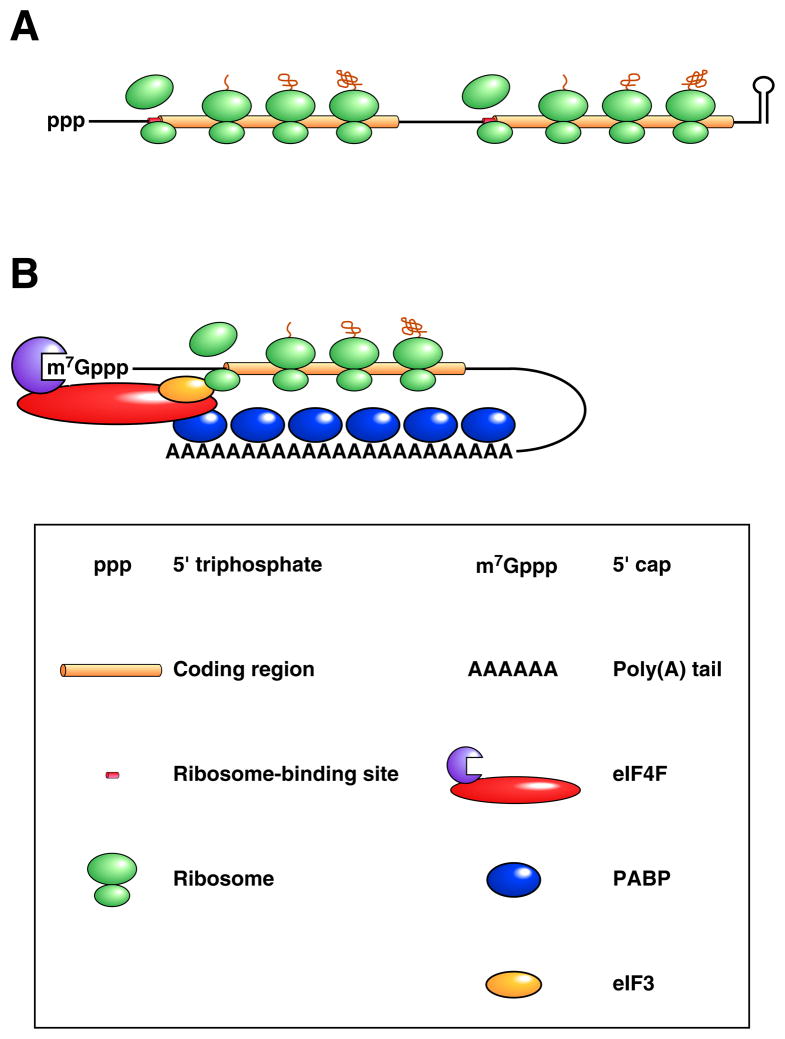
(A) Bacterial mRNA. A translated dicistronic transcript that begins with a triphosphate and ends with a stem-loop is depicted. Ribosome binding to the beginning of each translational unit is aided by base pairing between the 3’ end of 16S ribosomal RNA (a component of the small ribosomal subunit) and a Shine-Dalgarno element adjacent to the initiation codon (collectively termed the ribosome binding site). (B) Eukaryotic mRNA. A message undergoing cap-dependent translation is shown. Ribosome binding to the translation initiation codon is guided by the affinity of the small ribosomal subunit for eukaryotic initiation factor 3 (eIF3), a protein multimer recruited to mRNA by the cap-binding complex eIF4F, whose affinity for mRNA is enhanced by its ability to bind PABP associated with the 3’ poly(A) tail.
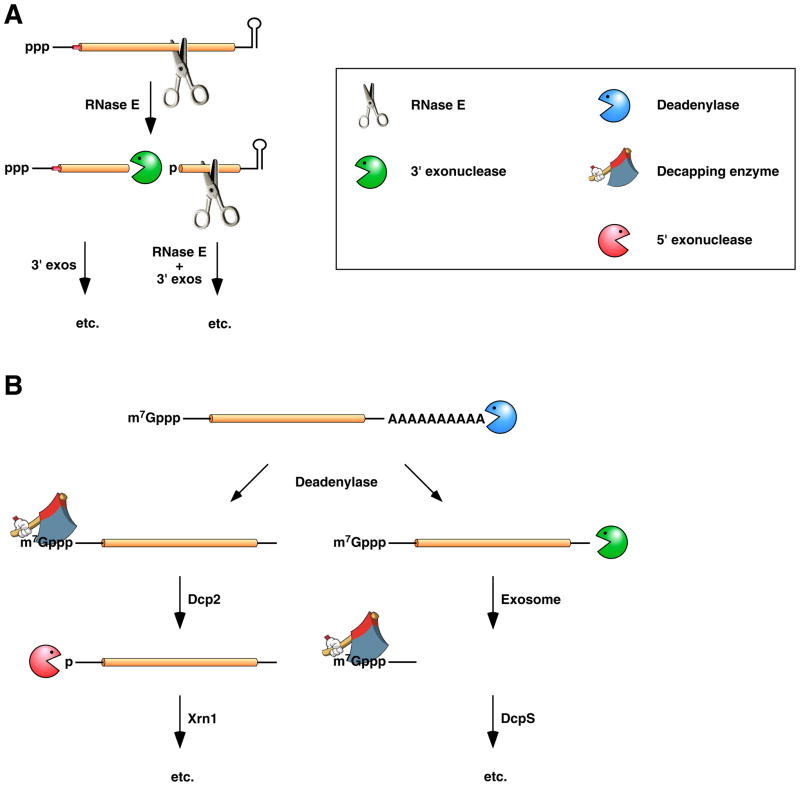
(A) mRNA decay in E. coli. In this pathway, serial internal cleavage by RNase E generates degradation intermediates whose lack of base pairing at the 3’ end renders them susceptible to attack by the 3’ exonucleases polynucleotide phosphorylase, RNase II, RNase R, and (for very short RNA fragments) oligoribonuclease. By contrast, the intact transcript resists exonucleolytic degradation because it is protected by a 3’-terminal stem-loop, which hinders such attack. (B) mRNA decay in eukaryotic cells. In this pathway, poly(A) tail removal by a deadenylase (Ccr4-Not, Pan2-Pan3, or PARN) yields a deadenylated intermediate susceptible both to decapping by Dcp2 and to 3’-exonucleolytic degradation by exosomes. The decapped RNA generated by Dcp2 is then degraded by the 5’-exonuclease Xrn1, whereas the 5’-terminal RNA fragment that results from extensive exosome digestion undergoes cap removal by an alternative decapping enzyme (DcpS) specific for oligonucleotides . These pathways were deduced from early studies of mRNA degradation in E. coli, S. cerevisiae, and mammalian cells. Ribosomes, PABP, and translation factors have been omitted from this figure for the sake of simplicity.
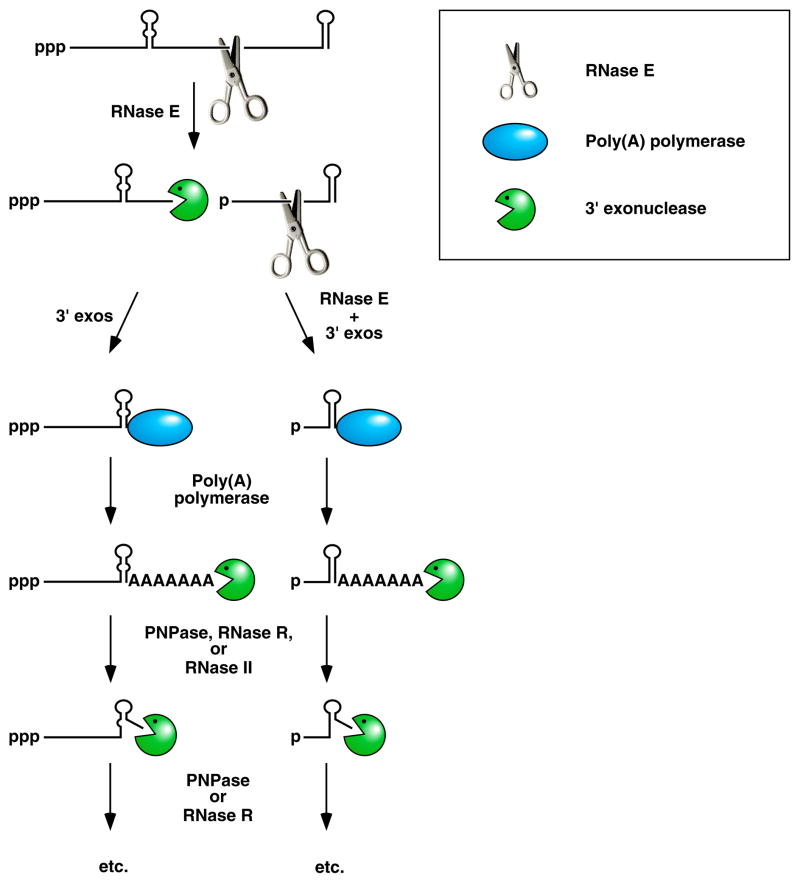
Endonucleolytic cleavage of mRNA by RNase E generates multiple fragments, one of which ends with the original 3’-terminal stem-loop. The others undergo 3’-exonucleolytic attack by PNPase, RNase R, and/or RNase II until an upstream stem-loop is encountered, which interrupts further degradation due to the preference of those ribonucleases for 3’ ends that are unpaired. The resulting decay intermediates are then polyadenylated by poly(A) polymerase, thereby enabling the exonucleases to re-engage. The repeated addition of single-stranded poly(A) tails to the 3’ ends of these intermediates provides multiple opportunities for PNPase and RNase R to overcome structural impediments to exonucleolytic degradation, and eventually they succeed. The ability of PNPase to digest base-paired RNA is enhanced by its association with the RNA helicase RhlB, whereas RNase R requires no such assistance. By contrast, RNase II can degrade poly(A) and other kinds of unstructured RNA but not structured RNA. Ribosomes and coding regions have been omitted from this figure for the sake of simplicity.
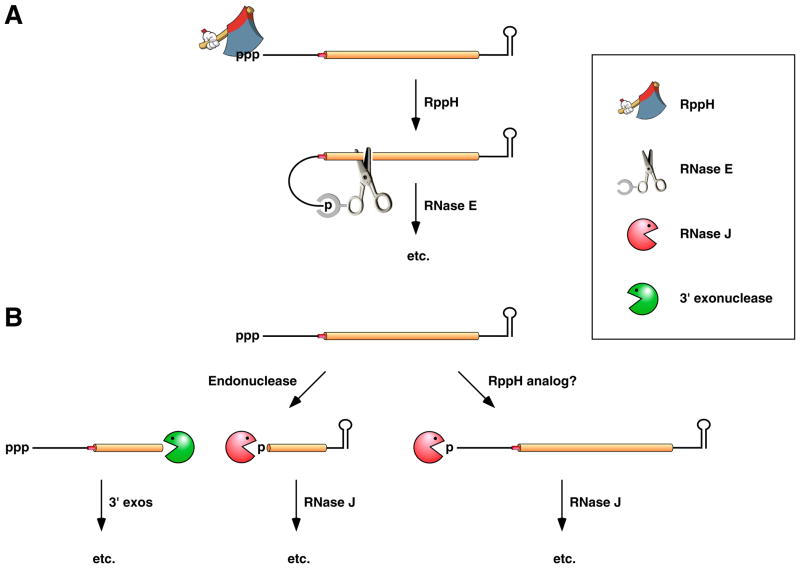
(A) 5’-end-dependent mRNA decay in bacteria that contain the endonuclease RNase E or a homolog thereof. Pyrophosphate removal by RppH generates a 5’-terminal monophosphate that binds to a discrete pocket on the surface of RNase E, thereby facilitating mRNA cleavage at a downstream location by the active site of that enzyme. In E. coli, RNase E cleavage of primary transcripts can also occur by an alternative, 5’-end-independent mechanism that does not require prior pyrophosphate removal (Figure 2A) , , . (B) 5’-end-dependent mRNA decay in bacteria that contain the 5’ exonuclease RNase J. Internal cleavage by an endonuclease generates a monophosphorylated intermediate susceptible to 5’-to-3’ digestion by RNase J, whose exonucleolytic activity is impeded by a 5’ triphosphate. Alternatively, it is possible that 5’-exonucleolytic digestion by RNase J may be triggered by pyrophosphate removal from primary transcripts by an as yet unidentified RppH analog. Ribosomes have been omitted from this figure for the sake of simplicity.
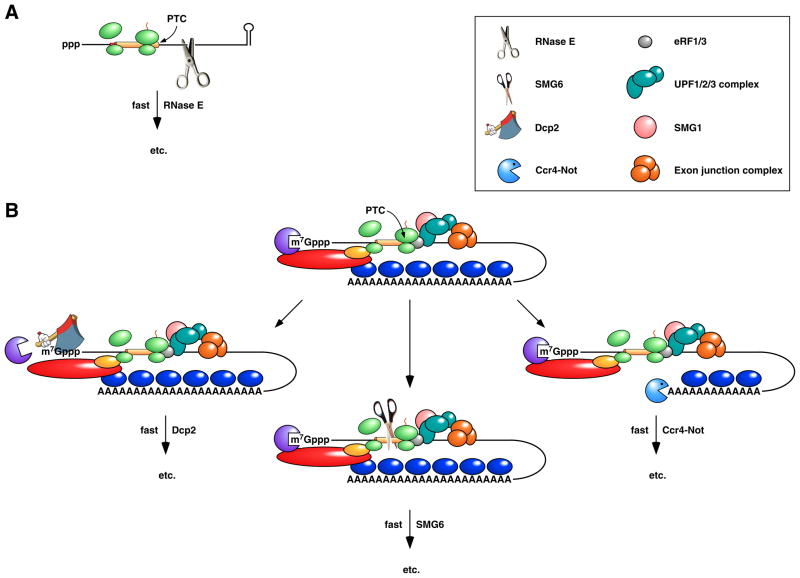
(A) Rapid degradation of a premature termination codon (PTC)-containing mRNA in E. coli. Premature translation termination and the resulting loss of ribosome protection downstream of the PTC expose the mRNA to internal cleavage by RNase E. (B) Nonsense-mediated decay (NMD) in metazoans. Premature translation termination results in an unusually long 3’ UTR, often in conjunction with an exon junction downstream of the PTC. The assembly of the PTC surveillance proteins UPF1, UPF2, and UPF3 at the site of translation termination is guided by the presence there of termination factors eRF1 and eRF3 and can be enhanced by the interaction of UPF3 with an exon junction complex, a heteromultimer deposited on exon junctions during splicing, transported with mRNA to the cytoplasm, and displaced by translating ribosomes only if bound in the coding region. Phosphorylation of UPF1 by the serine/threonine kinase SMG1 triggers nonsense-mediated decay via any of three pathways: deadenylation-independent decapping by Dcp2 (left), endonucleolytic cleavage by SMG6 (centre), or poly(A) tail removal by Ccr4-Not (right). Deadenylation-independent decapping or poly(A) removal leads to degradation via the pathways depicted in Figure 2B. Endonucleolytic cleavage leads to 5’-exonucleolytic degradation of the 3’ fragment by Xrn1 and degradation of the 5’ fragment via the pathways depicted in Figure 2B.
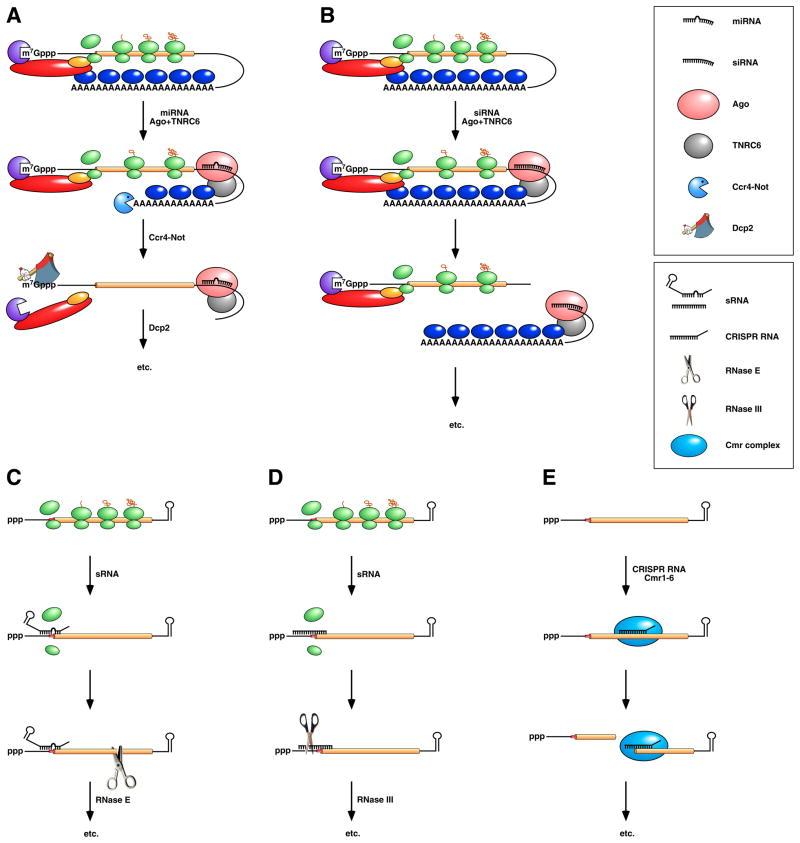
(Top) Repression by miRNAs in vertebrates and insects. (A) A miRNA associated with Ago and TNRC6 (also known as GW182) binds to the 3’ UTR of a message to which it is partially complementary, impeding (though not abolishing) translation and accelerating poly(A) tail removal by the deadenylase Ccr4-Not. The deadenylated mRNA is then decapped by Dcp2 and degraded exonucleolytically by Xrn1 and possibly also by exosomes. (B) An siRNA associated with Ago and TNRC6/GW182 binds to a message to which it is fully complementary, either within the 3’ UTR or somewhere upstream, and directs endonucleolytic cleavage there by Ago. Endonucleolytic cleavage leads to 5’-exonucleolytic degradation of the 3’ fragment by Xrn1 and degradation of the 5’ fragment via the pathways depicted in Figure 2B. Note that miRNAs and siRNAs have the same regulatory potential, their mode of action being determined by the degree of complementarity of their mRNA targets and the activity of the Ago and TNRC6/GW182 proteins with which they associate. (Bottom) Repression by sRNAs and CRISPR RNAs in bacteria. (C) sRNA binding to a partially complementary mRNA impairs translation initiation, often by occluding the ribosome binding site. No longer protected by ribosomes, the message becomes vulnerable to attack by RNase E. (D) Antisense sRNA binding to a fully complementary mRNA can create a long, perfectly paired duplex susceptible to cleavage by RNase III. (E) A Cmr-associated CRISPR RNA directs endonucleolytic cleavage of a complementary message by one of the six Cmr proteins. Ribosomes have been omitted from this panel because the effect of CRISPR RNAs on translation has not been investigated.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
