

Review
doi: 10.1038/nsmb0610-650.
The quest to understand heterotrimeric G protein signaling
Affiliations
- PMID: 20520658
- PMCID: PMC2954486
- DOI: 10.1038/nsmb0610-650
Item in Clipboard
Review
The quest to understand heterotrimeric G protein signaling
Nat Struct Mol Biol.
2010 Jun.
Abstract
Over the past fifteen years, great strides have been made to understand how heterotrimeric G proteins control their downstream targets. However, the mechanism by which heterotrimeric G proteins are activated by their upstream G protein-coupled receptors (GPCRs) remains obscure. Recent structural data support the idea that GPCRs, despite their small size, are sophisticated allosteric machines with multiple signaling outputs.
Figures
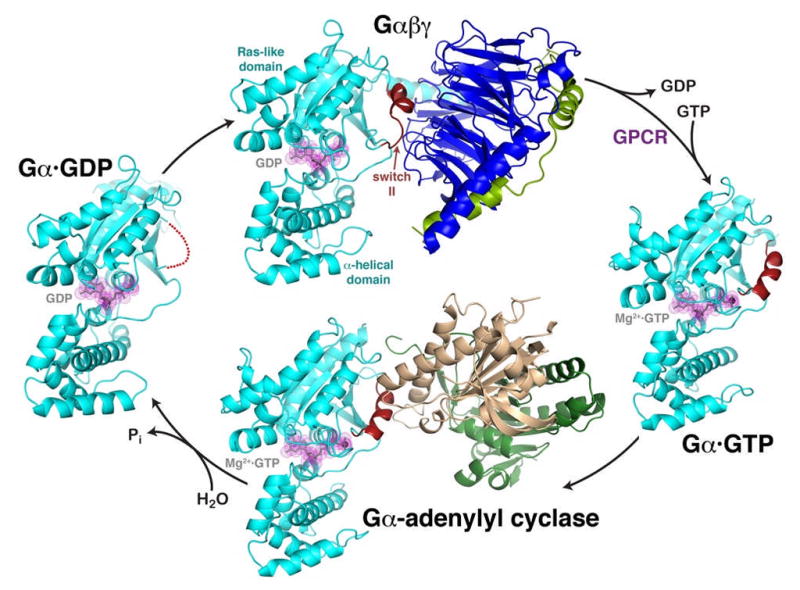
The heterotrimeric G protein cycle. The inactive Gαβγ heterotrimer (top) is composed of two principal elements, Gα·GDP (cyan, with GDP shown in grey with magenta dot surface) and the Gβγ heterodimer (blue and green). Gβγ sequesters the switch II element (red) such that it is unable to interact with effectors. Activated GPCRs catalyze the release of GDP from Gα, allowing GTP to bind and liberate the activated Gα·GTP subunit (right). In this state, switch II forms a helix stabilized by the γ-phosphate of GTP. The activated Gαsubunit can then interact with effectors, such as the catalytic domains of adenylyl cyclase (bottom, gold and dark green). Switch II forms a major component of the interface, as it does in other characterized effector complexes. The Gαsubunit has a slow GTPase activity that converts GTP to GDP, weakening its interactions with effectors and allowing it to dissociate as a deactivated Gα·GDP subunit (left). In this state, switch II is disordered (red dotted line), and the protein has high affinity for Gβγ subunits, completing the cycle. The structures shown correspond to PDB entries 1GG2 (ref. 27) (top, cyan subunit corresponds to Gαi), 1AZT28 (right, Gαs), 1AZS4 (bottom, Gαs) and 1GDD29 (left, Gαi).
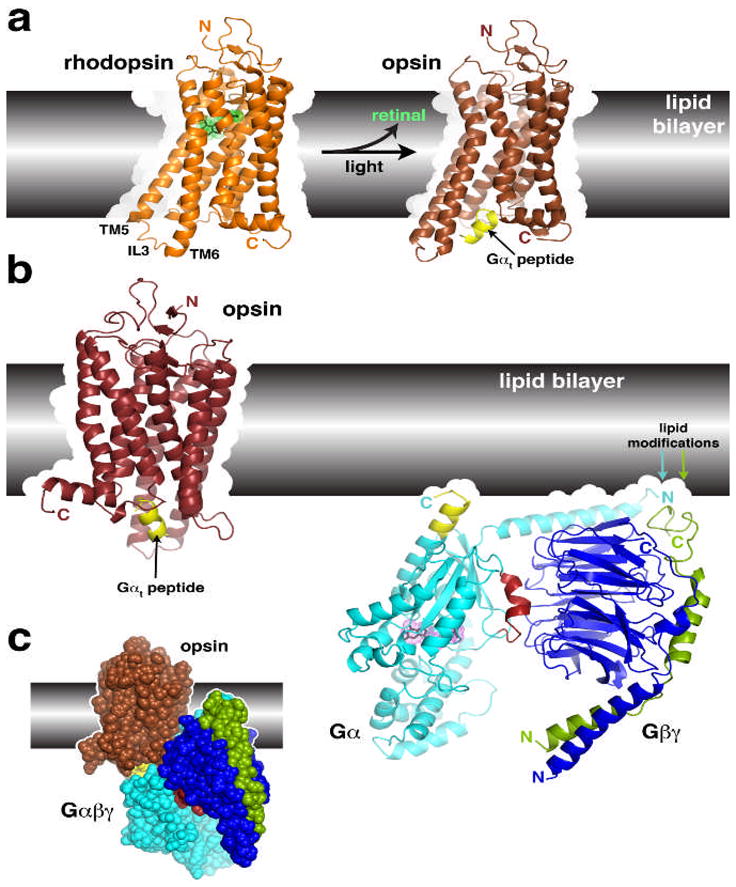
Structural transitions of GPCRs and their interactions with heterotrimeric G proteins. (a) Comparison of the structures of rhodopsin and opsin. Opsin represents a retinal-depleted form of the GPCR with low basal activity yet shows structural features consistent with changes predicted to occur during rhodopsin activation, such as the outward twist of the third intracellular loop (IL3). Opsin can also be crystallized in complex with a peptide derived from the C-terminus of Gαt (yellow), the region of the heterotrimer most strongly linked to receptor recognition. Retinal is shown as a stick model with a green dot surface. Structures correspond to PDB entries 1GZM 30 and 3DQB 17. (b) Conceptual problems in docking current models of GPCRs with their heterotrimeric G protein substrates. Here we show the inactive Gαiβ1γ2 complex (PDB 1GG2 (ref. 27)), in which the C-terminus of Gαi was extended according to PDB entry 1AGR31 and the C-terminus of Gγ according to PDB entry 1OMW32. The C-terminal span of Gαi analogous to the Gαt peptide bound to opsin is colored yellow. Comparing the orientation of this helix in each model reveals an an apparent docking incompatibility, because the intact heterotrimer would have to be rotated, roughly counterclockwise, up into the plane of the lipid bilayer in order to superimpose these elements. (c) Collision of Gαβγ with the lipid bilayer when superimposed with the Gαt peptide bound to opsin. The collision suggests that either the model of opsin does not represent a GPCR in a fully activated state, or the G protein heterotrimer must undergo a significant conformational change, or both. A large conformational change is expected in Gαβγ because its interaction with receptors must induce nucleotide release.
References
-
- Rall TW, Sutherland EW. The regulatory role of adenosine-3′, 5′-phosphate. Cold Spring Harb Symp Quant Biol. 1961;26:347–54. - PubMed
-
- Hepler JR, Gilman AG. G proteins. Trends Biochem Sci. 1992;17:383–7. - PubMed
-
- Sprang SR, Chen Z, Du X. Structural basis of effector regulation and signal termination in heterotrimeric G proteins. Adv Protein Chem. 2007;74:1–65. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
