

Understanding the role of PknJ in Mycobacterium tuberculosis: biochemical characterization and identification of novel substrate pyruvate kinase A
- PMID: 20520732
- PMCID: PMC2875399
- DOI: 10.1371/journal.pone.0010772
Understanding the role of PknJ in Mycobacterium tuberculosis: biochemical characterization and identification of novel substrate pyruvate kinase A
Abstract
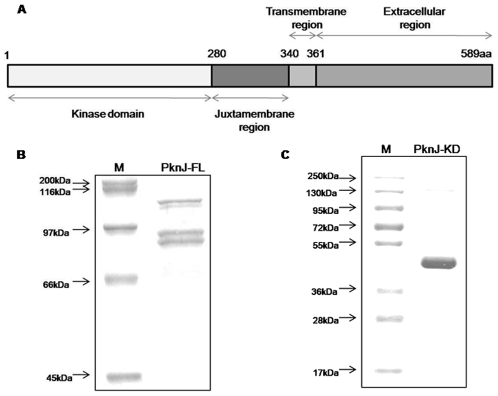
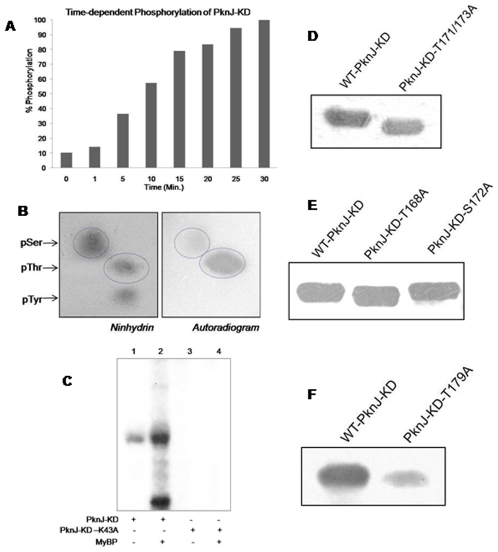
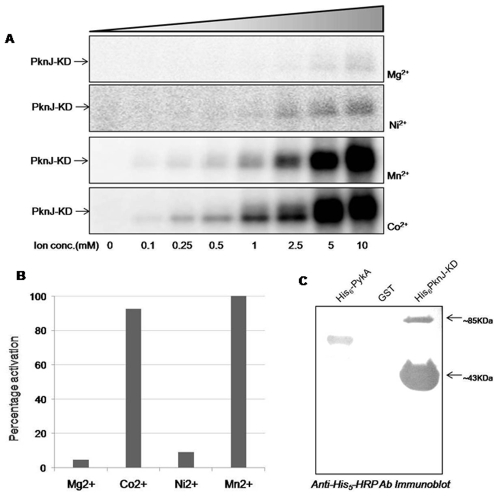
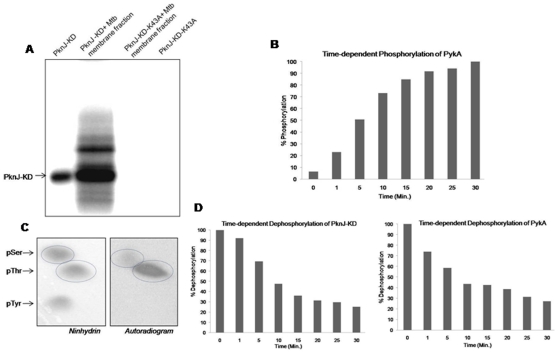
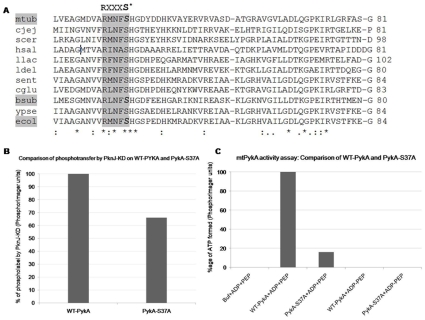
Reversible protein phosphorylation is a prevalent signaling mechanism which modulates cellular metabolism in response to changing environmental conditions. In this study, we focus on previously uncharacterized Mycobacterium tuberculosis Ser/Thr protein kinase (STPK) PknJ, a putative transmembrane protein. PknJ is shown to possess autophosphorylation activity and is also found to be capable of carrying out phosphorylation on the artificial substrate myelin basic protein (MyBP). Previous studies have shown that the autophosphorylation activity of M. tuberculosis STPKs is dependent on the conserved residues in the activation loop. However, our results show that apart from the conventional conserved residues, additional residues in the activation loop may also play a crucial role in kinase activation. Further characterization of PknJ reveals that the kinase utilizes unusual ions (Ni(2+), Co(2+)) as cofactors, thus hinting at a novel mechanism for PknJ activation. Additionally, as shown for other STPKs, we observe that PknJ possesses the capability to dimerize. In order to elucidate the signal transduction cascade emanating from PknJ, the M. tuberculosis membrane-associated protein fraction is treated with the active kinase and glycolytic enzyme Pyruvate kinase A (mtPykA) is identified as one of the potential substrates of PknJ. The phospholabel is found to be localized on serine and threonine residue(s), with Ser(37) identified as one of the sites of phosphorylation. Since Pyk is known to catalyze the last step of glycolysis, our study shows that the fundamental pathways such as glycolysis can also be governed by STPK-mediated signaling.
Conflict of interest statement
Figures
References
-
- Cole ST, Brosch R, Parkhill J, Garnier T, Churcher C, et al. Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature. 1998;393:537–544. - PubMed
-
- Bach H, Wong D, Av-Gay Y. Mycobacterium tuberculosis PtkA is a novel protein tyrosine kinase whose substrate is PtpA. Biochem J. 2009;420:155–160. - PubMed
-
- Vohra R, Gupta M, Chaturvedi R, Singh Y. Attack on the scourge of Tuberculosis: Patented Drug Targets. Recent Pat Antiinfect Drug Discov. 2006;1:95–106. - PubMed
-
- Chaba R, Raje M, Chakraborti PK. Evidence that a eukaryotic-type serine/threonine protein kinase from Mycobacterium tuberculosis regulates morphological changes associated with cell division. Eur J Biochem. 2002;269:1078–1085. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
