

A mutant allele of the Swi/Snf member BAF250a determines the pool size of fetal liver hemopoietic stem cell populations
- PMID: 20522713
- PMCID: PMC2947392
- DOI: 10.1182/blood-2010-03-273862
A mutant allele of the Swi/Snf member BAF250a determines the pool size of fetal liver hemopoietic stem cell populations
Abstract
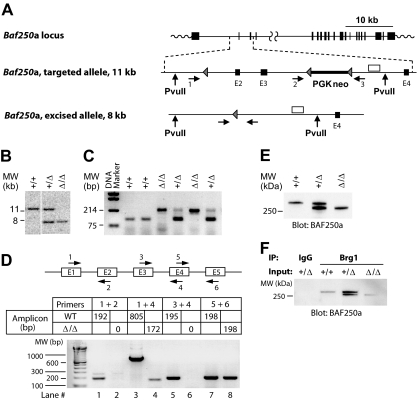
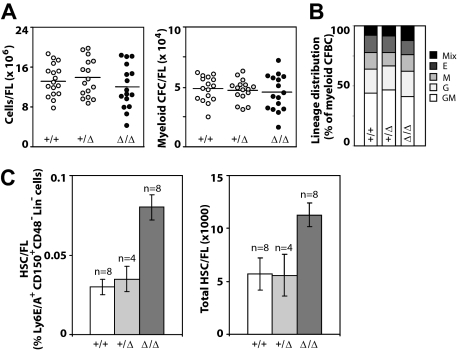
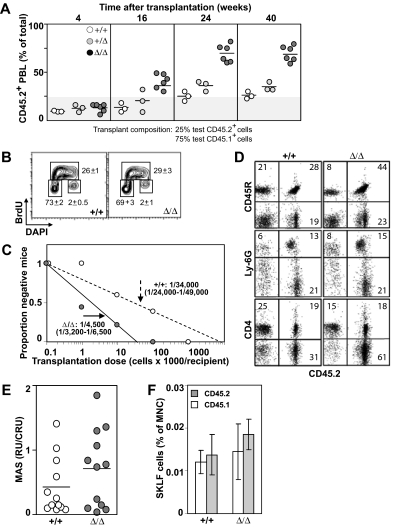
It is believed that hemopoietic stem cells (HSC), which colonize the fetal liver (FL) rapidly, expand to establish a supply of HSCs adequate for maintenance of hemopoiesis throughout life. Accordingly, FL HSCs are actively cycling as opposed to their predominantly quiescent bone marrow counterparts, suggesting that the FL microenvironment provides unique signals that support HSC proliferation and self-renewal. We now report the generation and characterization of mice with a mutant allele of Baf250a lacking exons 2 and 3. Baf250a(E2E3/E2E3) mice are viable until E19.5, but do not survive beyond birth. Most interestingly, FL HSC numbers are markedly higher in these mice than in control littermates, thus raising the possibility that Baf250a determines the HSC pool size in vivo. Limit dilution experiments indicate that the activity of Baf250a(E2E3/E2E3) HSC is equivalent to that of the wild-type counterparts. The Baf250a(E2E3/E2E3) FL-derived stroma, in contrast, exhibits a hemopoiesis-supporting potential superior to the developmentally matched controls. To our knowledge, this demonstration is the first that a mechanism operating in a cell nonautonomous manner canexpand the pool size of the fetal HSC populations.
Figures

References
-
- Mikkola HK, Orkin SH. The journey of developing hematopoietic stem cells. Development. 2006;133(19):3733–3744. - PubMed
-
- Ema H, Nakauchi H. Expansion of hematopoietic stem cells in the developing liver of a mouse embryo. Blood. 2000;95(7):2284–2288. - PubMed
-
- Wilson A, Laurenti E, Oser G, et al. Hematopoietic stem cells reversibly switch from dormancy to self-renewal during homeostasis and repair. Cell. 2008;135(6):1118–1129. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
