

The landscape of C. elegans 3'UTRs
- PMID: 20522740
- PMCID: PMC3142571
- DOI: 10.1126/science.1191244
The landscape of C. elegans 3'UTRs
Abstract
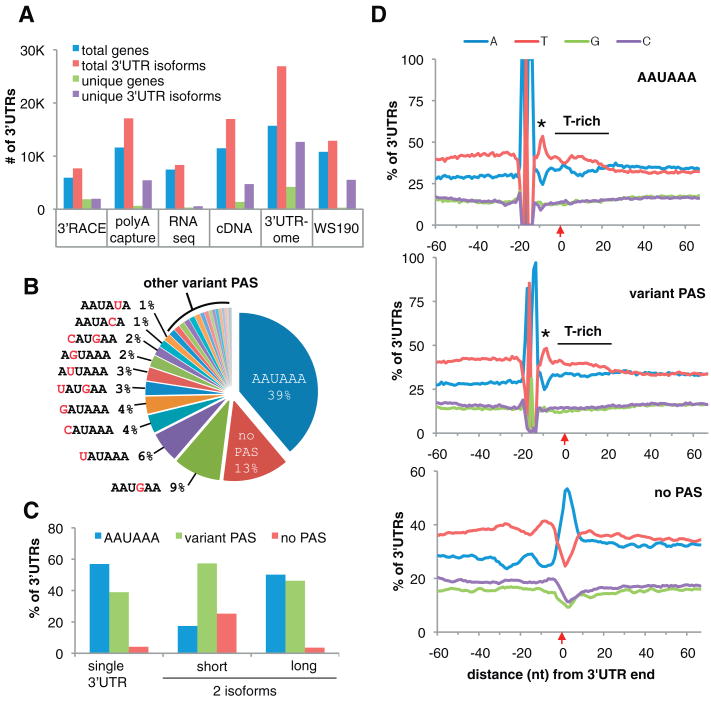
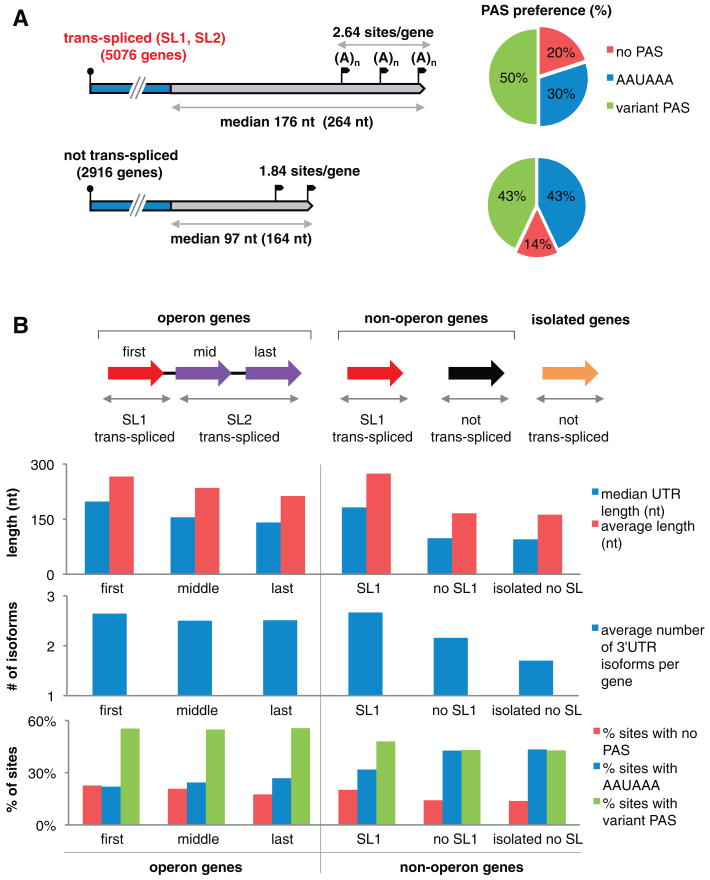
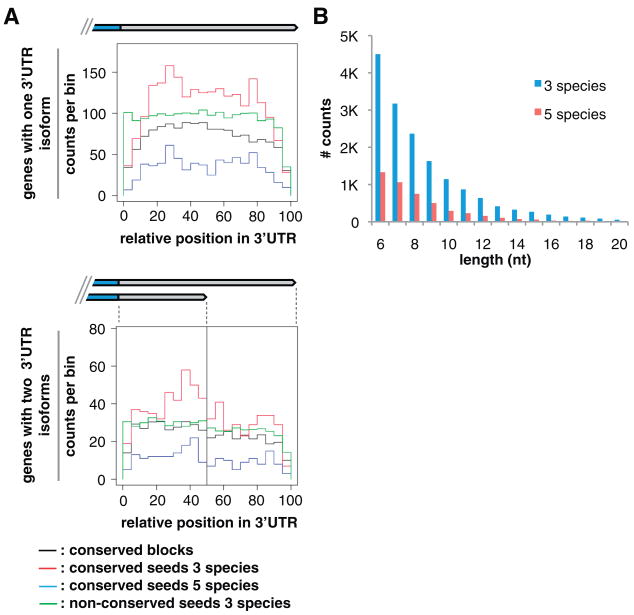
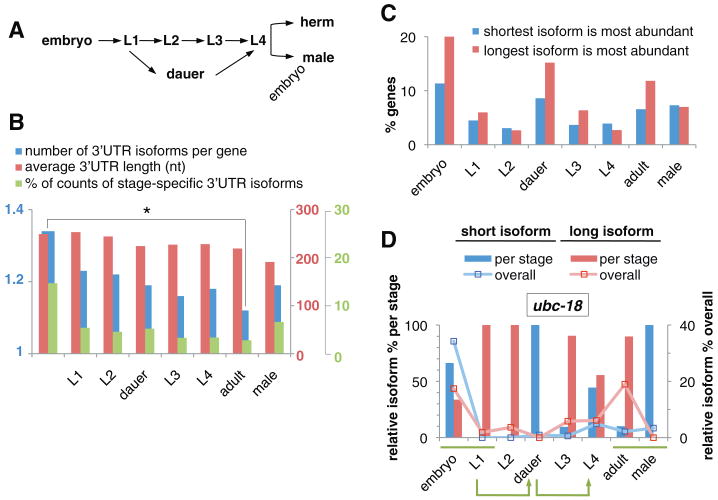
Three-prime untranslated regions (3'UTRs) of metazoan messenger RNAs (mRNAs) contain numerous regulatory elements, yet remain largely uncharacterized. Using polyA capture, 3' rapid amplification of complementary DNA (cDNA) ends, full-length cDNAs, and RNA-seq, we defined approximately 26,000 distinct 3'UTRs in Caenorhabditis elegans for approximately 85% of the 18,328 experimentally supported protein-coding genes and revised approximately 40% of gene models. Alternative 3'UTR isoforms are frequent, often differentially expressed during development. Average 3'UTR length decreases with animal age. Surprisingly, no polyadenylation signal (PAS) was detected for 13% of polyadenylation sites, predominantly among shorter alternative isoforms. Trans-spliced (versus non-trans-spliced) mRNAs possess longer 3'UTRs and frequently contain no PAS or variant PAS. We identified conserved 3'UTR motifs, isoform-specific predicted microRNA target sites, and polyadenylation of most histone genes. Our data reveal a rich complexity of 3'UTRs, both genome-wide and throughout development.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
