

Preferential oxidation of triacylglyceride-derived fatty acids in heart is augmented by the nuclear receptor PPARalpha
- PMID: 20522803
- PMCID: PMC2921193
- DOI: 10.1161/CIRCRESAHA.110.221713
Preferential oxidation of triacylglyceride-derived fatty acids in heart is augmented by the nuclear receptor PPARalpha
Abstract
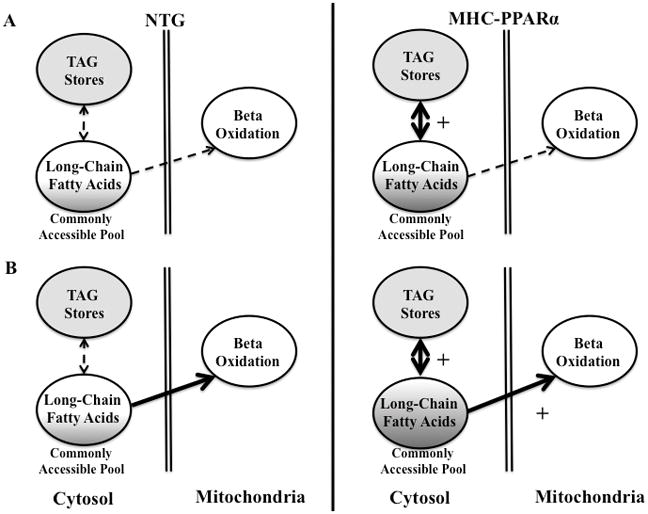
Rationale: Long chain fatty acids (LCFAs) are the preferred substrate for energy provision in hearts. However, the contribution of endogenous triacylglyceride (TAG) turnover to LCFA oxidation and the overall dependence of mitochondrial oxidation on endogenous lipid is largely unstudied.
Objective: We sought to determine the role of TAG turnover in supporting LCFA oxidation and the influence of the lipid-activated nuclear receptor, proliferator-activated receptor (PPAR)alpha, on this balance.
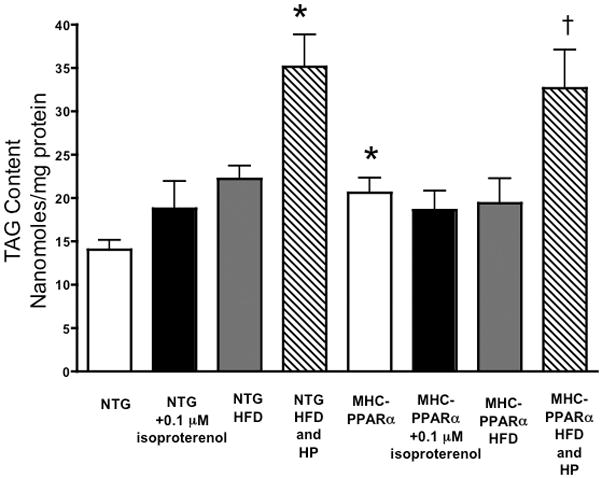
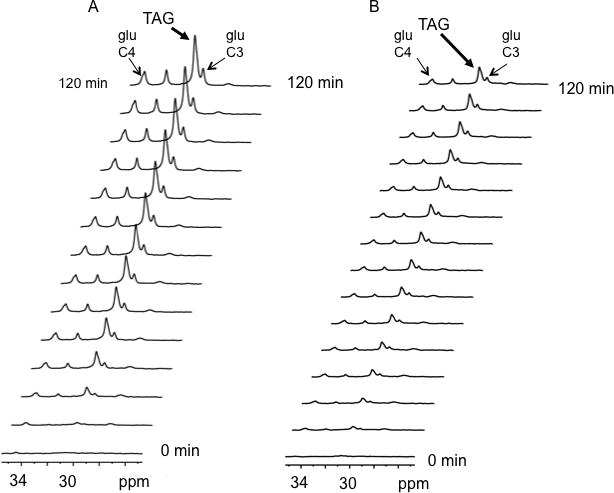
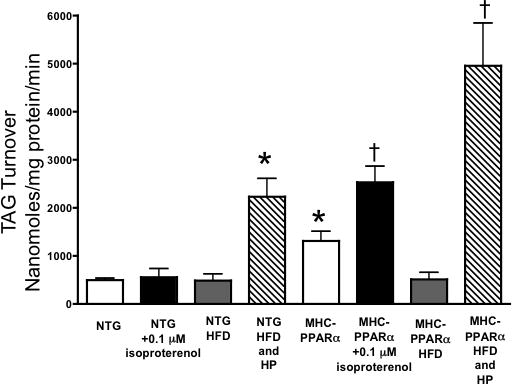
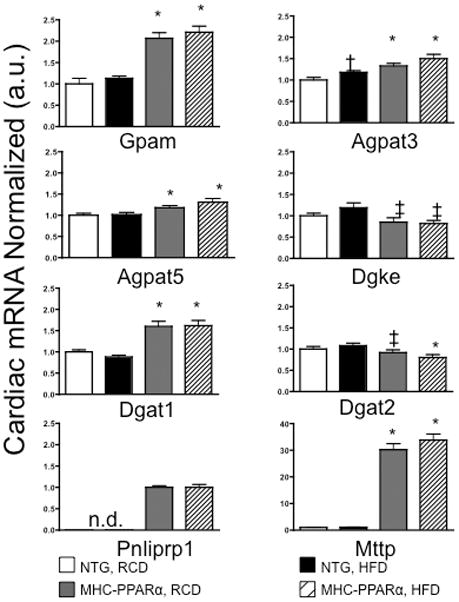
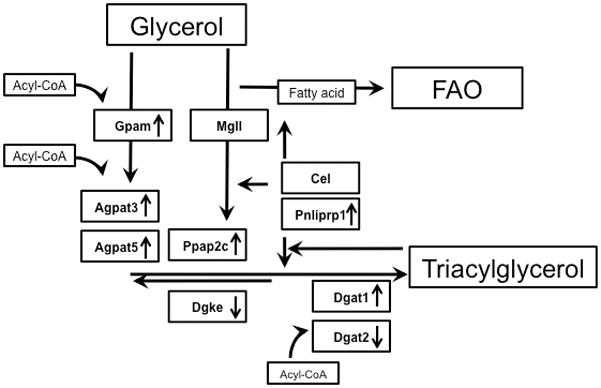
Methods and results: Palmitoyl turnover within TAG and palmitate oxidation rates were quantified in isolated hearts, from normal mice (nontransgenic) and mice with cardiac-specific overexpression of PPARalpha (MHC-PPARalpha). Turnover of palmitoyl units within TAG, and thus palmitoyl-coenzyme A recycling, in nontransgenic (4.5+/-2.3 micromol/min per gram dry weight) was 3.75-fold faster than palmitate oxidation (1.2+/-0.4). This high rate of palmitoyl unit turnover indicates preferential oxidation of palmitoyl units derived from TAG in normal hearts. PPARalpha overexpression augmented TAG turnover 3-fold over nontransgenic hearts, despite similar fractions of acetyl-coenzyme A synthesis from palmitate and oxygen use at the same workload. Palmitoyl turnover within TAG of MHC-PPARalpha hearts (16.2+/-2.9, P<0.05) was 12.5-fold faster than oxidation (1.3+/-0.2). Elevated TAG turnover in MHC-PPARalpha correlated with increased mRNA for enzymes involved in both TAG synthesis, Gpam (glycerol-3-phosphate acyltransferase, mitochondrial), Dgat1 (diacylglycerol acetyltransferase 1), and Agpat3 (1-acylglycerol-3-phospate O-acyltransferase 3), and lipolysis, Pnliprp1 (pancreatic lipase related protein 1).
Conclusions: The role of endogenous TAG in supporting beta-oxidation in the normal heart is much more dynamic than previously thought, and lipolysis provides the bulk of LCFA for oxidation. Accelerated palmitoyl turnover in TAG, attributable to chronic PPARalpha activation, results in near requisite oxidation of LCFAs from TAG.
Figures

References
-
- Neely JR, Morgan HE. Relationship between carbohydrate and lipid metabolism and the energy balance of heart muscle. Annu Rev Physiol. 1974;36:413–59. - PubMed
-
- Lopaschuk GD, Collins-Nakai RL, Itoi T. Developmental changes in energy substrate use by the heart. Cardiovasc Res. 1997;26:1172–80. - PubMed
-
- Russell LK, Finck BN, Kelly DP. Mouse models of mitochondrial dysfunction and heart failure. J Mol Cell Cardiol. 2005;38:81–91. - PubMed
-
- Finck BN. The role of the peroxisome proliferators-activated receptor alpha pathway in pathological remodeling of the diabetic heart. Curr Opin Clin Nutr Metab Care. 2004;7:391–6. - PubMed
-
- Sambandam N, Morabito D, Wagg C, Finck BN, Kelly DP, Lopaschuk GD. Chronic activation of PPARαis detrimental to cardiac recovery of ischemia. Am J Physiol Heart Circ Physiol. 2006;290:H87–95. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
