

miR-22 forms a regulatory loop in PTEN/AKT pathway and modulates signaling kinetics
- PMID: 20523723
- PMCID: PMC2877705
- DOI: 10.1371/journal.pone.0010859
miR-22 forms a regulatory loop in PTEN/AKT pathway and modulates signaling kinetics
Abstract
Background: The tumor suppressor PTEN (phosphatase and tensin homolog) is a lipid phosphatase that converts PIP3 into PIP2 and downregulates the kinase AKT and its proliferative and anti-apoptotic activities. The FoxO transcription factors are PTEN downstream effectors whose activity is negatively regulated by AKT-mediated phosphorylation. PTEN activity is frequently lost in many types of cancer, leading to increased cell survival and cell cycle progression.
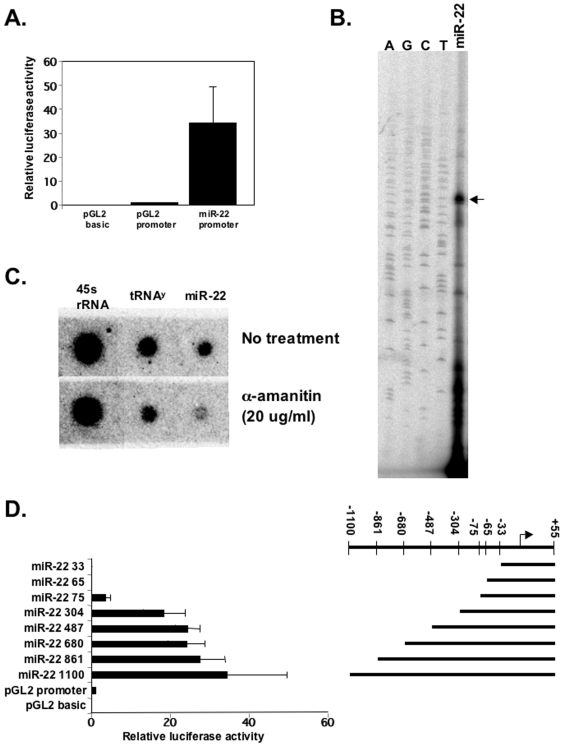
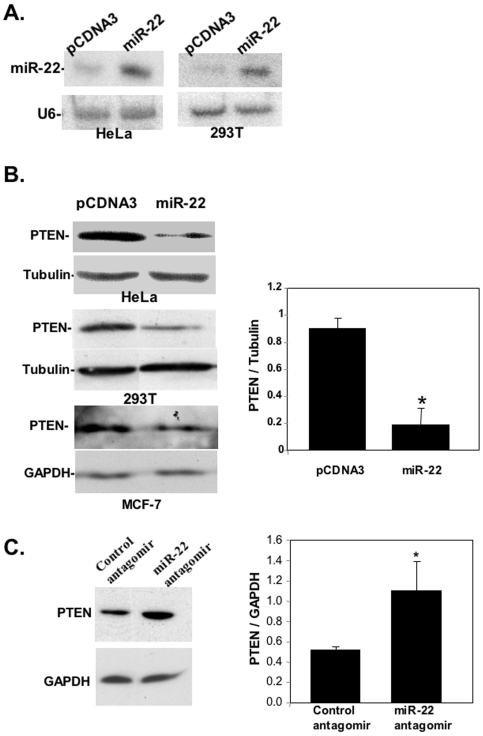
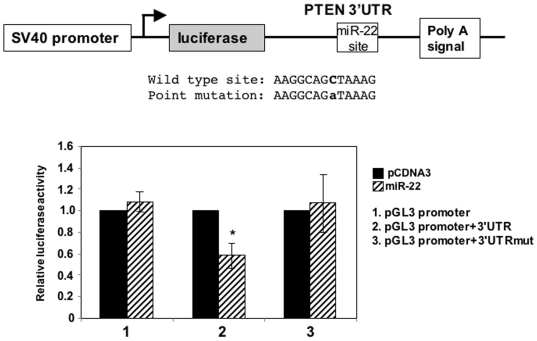
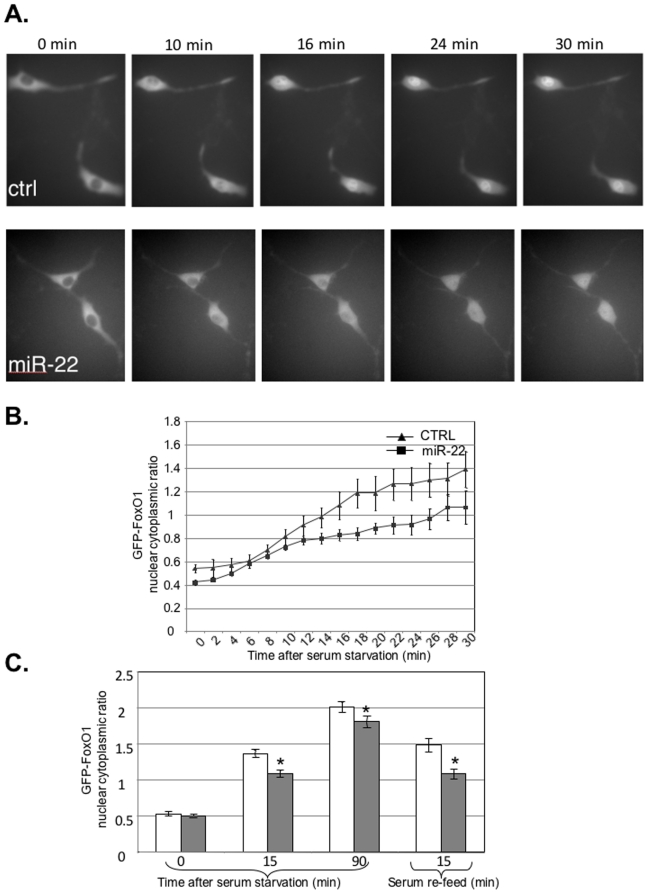
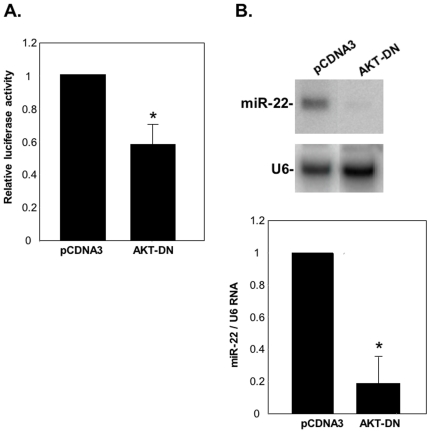
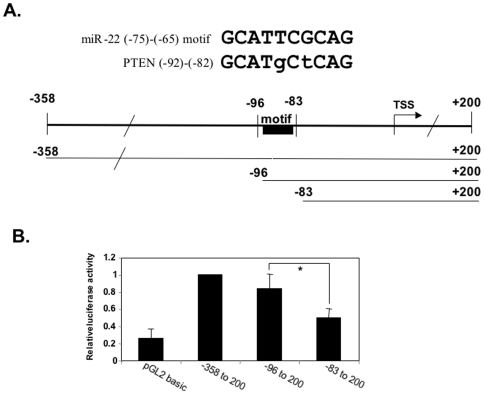
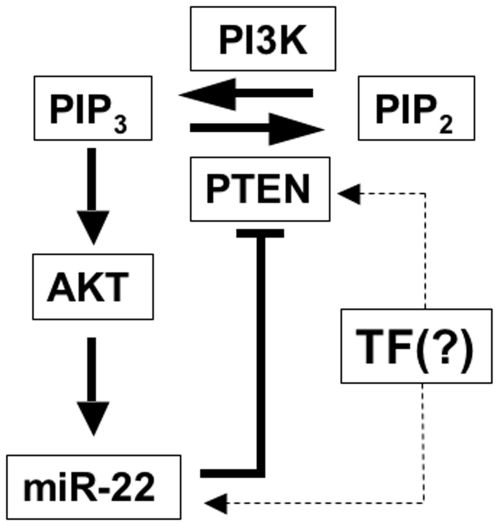
Principal findings: Here we characterize the widely expressed miR-22 and report that miR-22 is a novel regulatory molecule in the PTEN/AKT pathway. miR-22 downregulates PTEN levels acting directly through a specific site on PTEN 3'UTR. Interestingly, miR-22 itself is upregulated by AKT, suggesting that miR-22 forms a feed-forward circuit in this pathway. Time-resolved live imaging of AKT-dependent FoxO1 phosphorylation revealed that miR-22 accelerated AKT activity upon growth factor stimulation, and attenuated its down regulation by serum withdrawal.
Conclusions: Our results suggest that miR-22 acts to fine-tune the dynamics of PTEN/AKT/FoxO1 pathway.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
