

Competition of Escherichia coli DNA polymerases I, II and III with DNA Pol IV in stressed cells
- PMID: 20523737
- PMCID: PMC2877720
- DOI: 10.1371/journal.pone.0010862
Competition of Escherichia coli DNA polymerases I, II and III with DNA Pol IV in stressed cells
Abstract
Escherichia coli has five DNA polymerases, one of which, the low-fidelity Pol IV or DinB, is required for stress-induced mutagenesis in the well-studied Lac frameshift-reversion assay. Although normally present at approximately 200 molecules per cell, Pol IV is recruited to acts of DNA double-strand-break repair, and causes mutagenesis, only when at least two cellular stress responses are activated: the SOS DNA-damage response, which upregulates DinB approximately 10-fold, and the RpoS-controlled general-stress response, which upregulates Pol IV about 2-fold. DNA Pol III was also implicated but its role in mutagenesis was unclear. We sought in vivo evidence on the presence and interactions of multiple DNA polymerases during stress-induced mutagenesis. Using multiply mutant strains, we provide evidence of competition of DNA Pols I, II and III with Pol IV, implying that they are all present at sites of stress-induced mutagenesis. Previous data indicate that Pol V is also present. We show that the interactions of Pols I, II and III with Pol IV result neither from, first, induction of the SOS response when particular DNA polymerases are removed, nor second, from proofreading of DNA Pol IV errors by the editing functions of Pol I or Pol III. Third, we provide evidence that Pol III itself does not assist with but rather inhibits Pol IV-dependent mutagenesis. The data support the remaining hypothesis that during the acts of DNA double-strand-break (DSB) repair, shown previously to underlie stress-induced mutagenesis in the Lac system, there is competition of DNA polymerases I, II and III with DNA Pol IV for action at the primer terminus. Up-regulation of Pol IV, and possibly other stress-response-controlled factor(s), tilt the competition in favor of error-prone Pol IV at the expense of more accurate polymerases, thus producing stress-induced mutations. This mutagenesis assay reveals the DNA polymerases operating in DSB repair during stress and also provides a sensitive indicator for DNA polymerase competition and choice in vivo.
Conflict of interest statement
Figures
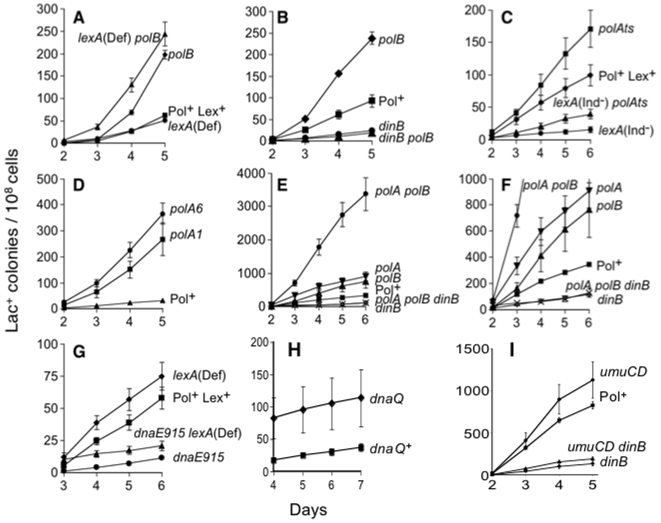

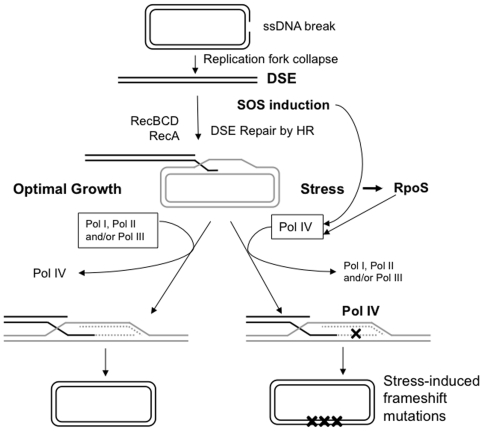
References
-
- Friedberg EC, Walker GC, Siede W, Wood RD, Schultz RA, et al. Washington, D. C.: ASM Press; 2005. DNA Repair and Mutagenesis.
-
- Kim S-Y, Maenhaut-Michel G, Yamada M, Yamamoto Y, Matsui K, et al. Multiple pathways for SOS-induced mutagenesis in Escherichia coli: an SOS gene product (DinB/P) enhances frameshift mutations in the absence of any exogenous agents that damage DNA. Proc Natl Acad Sci U S A. 1997;94:13792–13797. - PMC - PubMed
-
- Sommer S, Knezevic J, Bailone A, Devoret R. Induction of only one SOS operon, umuDC, is required for SOS mutagenesis in Escherichia coli. Mol Gen Genet. 1993;239:137–144. - PubMed
-
- Ohmori H, Friedberg EC, Fuchs RPP, Goodman MF, Hanaoka F, et al. The Y-family of DNA polymerases. Mol Cell. 2001;8:7–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
