

NK cells and gammadelta T cells mediate resistance to polyomavirus-induced tumors
- PMID: 20523894
- PMCID: PMC2877738
- DOI: 10.1371/journal.ppat.1000924
NK cells and gammadelta T cells mediate resistance to polyomavirus-induced tumors
Abstract
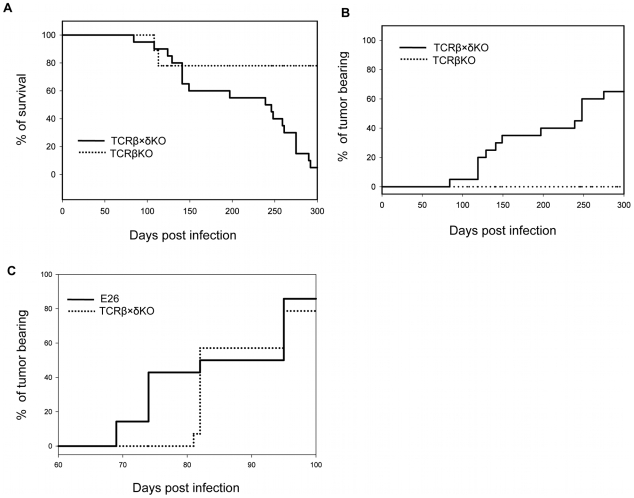
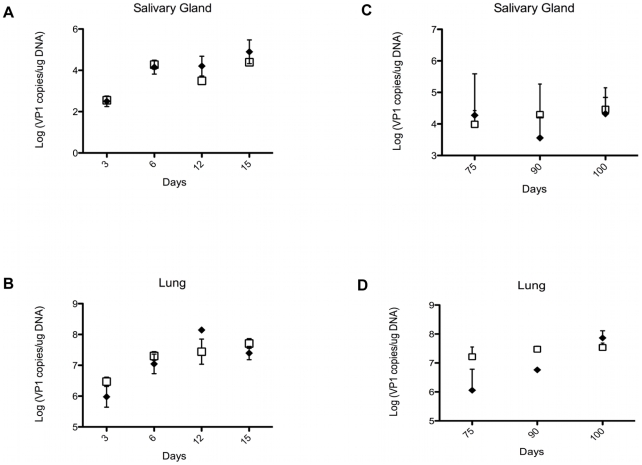
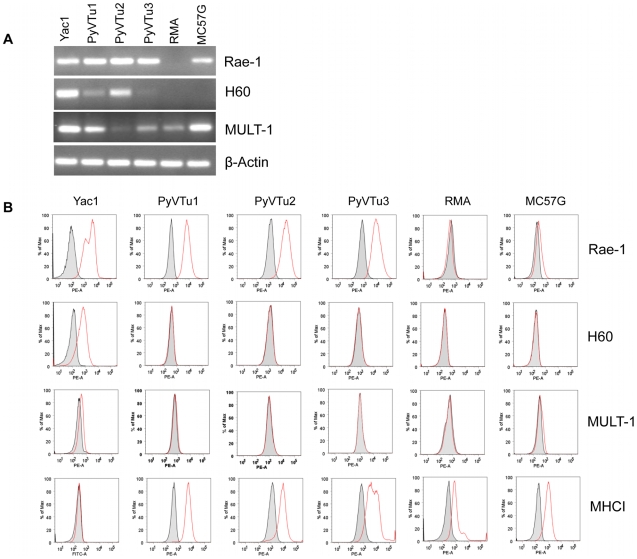
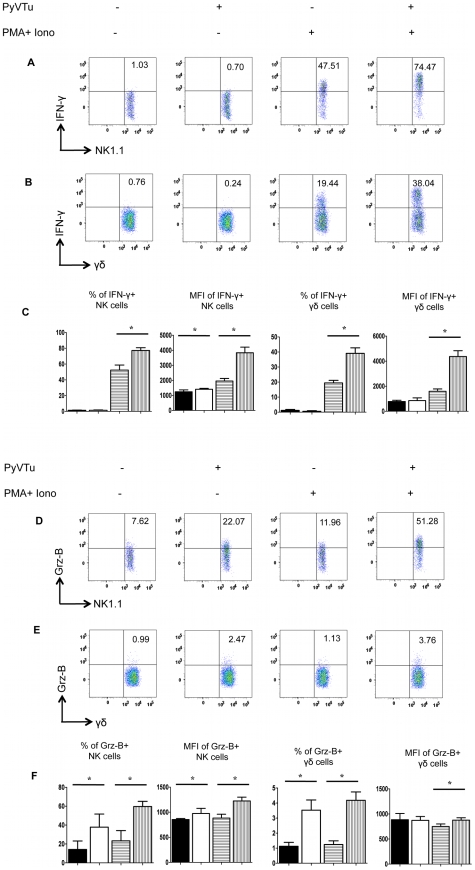
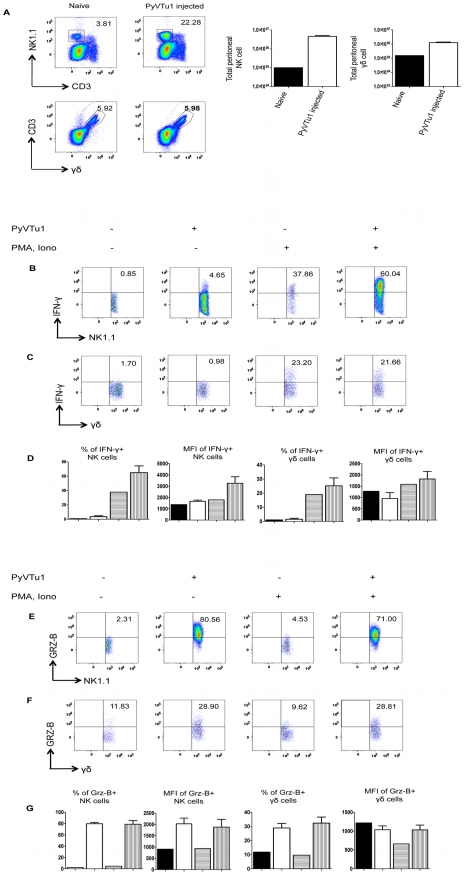
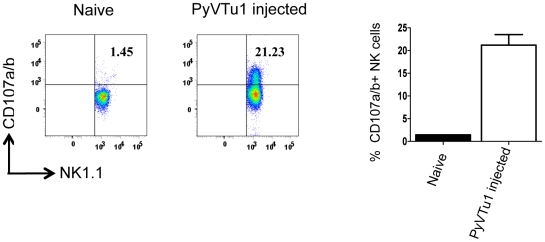
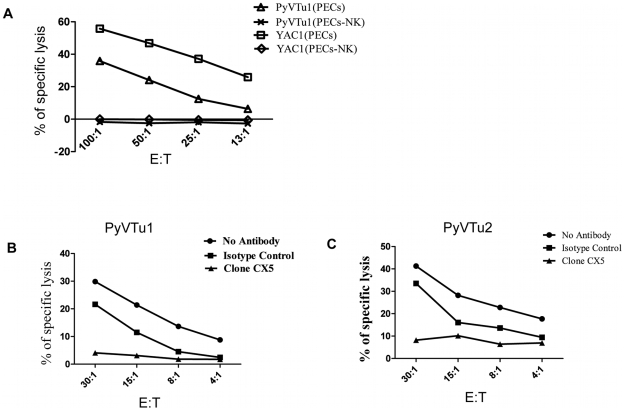
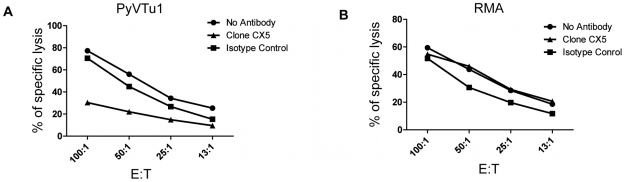
NK and gammadelta T cells can eliminate tumor cells in many experimental models, but their effect on the development of tumors caused by virus infections in vivo is not known. Polyomavirus (PyV) induces tumors in neonatally infected mice of susceptible strains and in adult mice with certain immune deficiencies, and CD8+ alphabeta T cells are regarded as the main effectors in anti-tumor immunity. Here we report that adult TCRbeta knockout (KO) mice that lack alphabeta but have gammadelta T cells remain tumor-free after PyV infection, whereas TCRbeta x delta KO mice that lack all T cells develop tumors. In addition, E26 mice, which lack NK and T cells, develop the tumors earlier than TCRbeta x delta KO mice. These observations implicate gammadelta T and NK cells in the resistance to PyV-induced tumors. Cell lines established from PyV-induced tumors activate NK and gammadelta T cells both in culture and in vivo and express Rae-1, an NKG2D ligand. Moreover, these PyV tumor cells are killed by NK cells in vitro, and this cytotoxicity is prevented by treatment with NKG2D-blocking antibodies. Our findings demonstrate a protective role for NK and gammadelta T cells against naturally occurring virus-induced tumors and suggest the involvement of NKG2D-mediated mechanisms.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Corvaisier M, Moreau-Aubry A, Diez E, Bennouna J, Mosnier JF, et al. V gamma 9V delta 2 T cell response to colon carcinoma cells. J Immunol. 2005;175:5481–5488. - PubMed
-
- Todaro M, D'Asaro M, Caccamo N, Iovino F, Francipane MG, et al. Efficient killing of human colon cancer stem cells by gammadelta T lymphocytes. J Immunol. 2009;182:7287–7296. - PubMed
-
- Takanami I, Takeuchi K, Giga M. The prognostic value of natural killer cell infiltration in resected pulmonary adenocarcinoma. J Thorac Cardiovasc Surg. 2001;121:1058–1063. - PubMed
-
- Coca S, Perez-Piqueras J, Martinez D, Colmenarejo A, Saez MA, et al. The prognostic significance of intratumoral natural killer cells in patients with colorectal carcinoma. Cancer. 1997;79:2320–2328. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
