

Effective, broad spectrum control of virulent bacterial infections using cationic DNA liposome complexes combined with bacterial antigens
- PMID: 20523903
- PMCID: PMC2877747
- DOI: 10.1371/journal.ppat.1000921
Effective, broad spectrum control of virulent bacterial infections using cationic DNA liposome complexes combined with bacterial antigens
Abstract
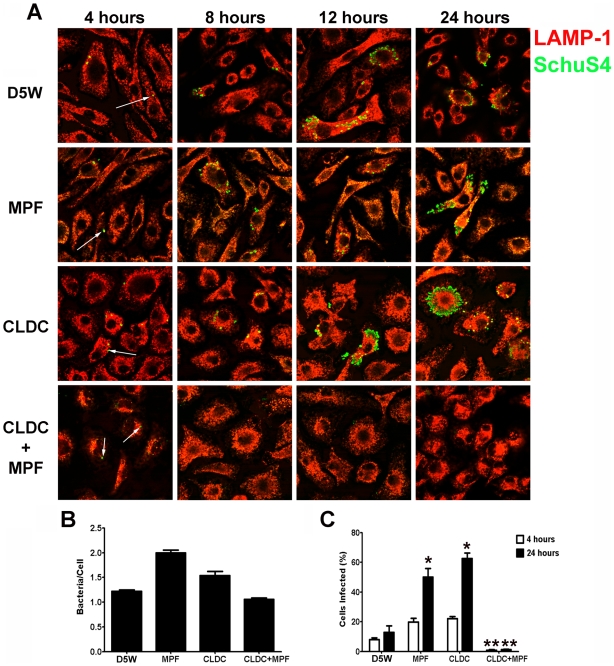
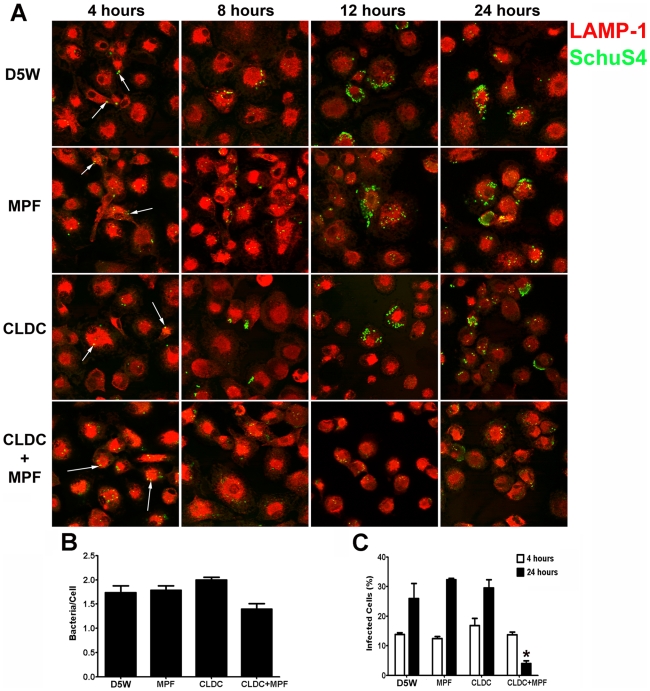
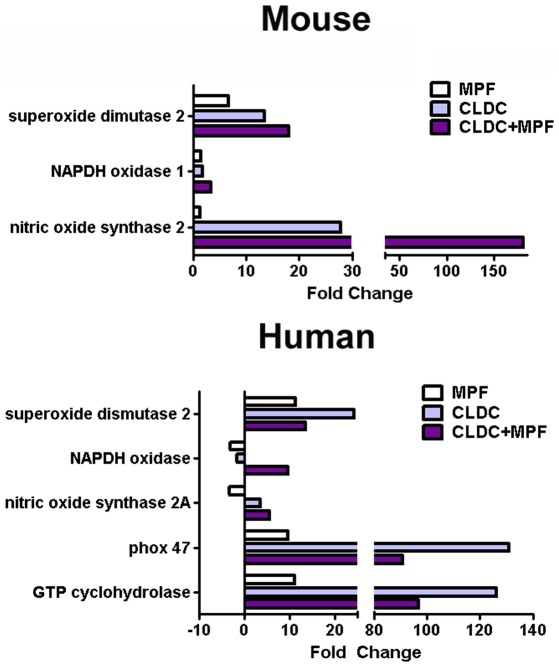
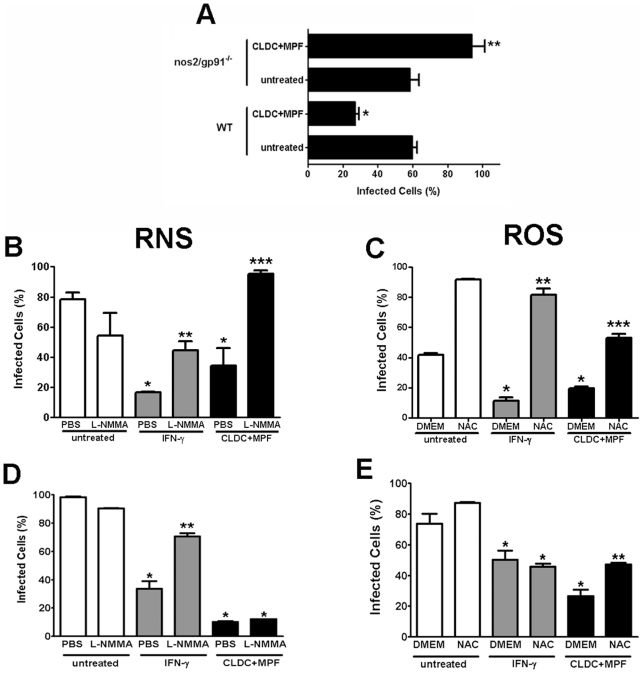
Protection against virulent pathogens that cause acute, fatal disease is often hampered by development of microbial resistance to traditional chemotherapeutics. Further, most successful pathogens possess an array of immune evasion strategies to avoid detection and elimination by the host. Development of novel, immunomodulatory prophylaxes that target the host immune system, rather than the invading microbe, could serve as effective alternatives to traditional chemotherapies. Here we describe the development and mechanism of a novel pan-anti-bacterial prophylaxis. Using cationic liposome non-coding DNA complexes (CLDC) mixed with crude F. tularensis membrane protein fractions (MPF), we demonstrate control of virulent F. tularensis infection in vitro and in vivo. CLDC+MPF inhibited bacterial replication in primary human and murine macrophages in vitro. Control of infection in macrophages was mediated by both reactive nitrogen species (RNS) and reactive oxygen species (ROS) in mouse cells, and ROS in human cells. Importantly, mice treated with CLDC+MPF 3 days prior to challenge survived lethal intranasal infection with virulent F. tularensis. Similarly to in vitro observations, in vivo protection was dependent on the presence of RNS and ROS. Lastly, CLDC+MPF was also effective at controlling infections with Yersinia pestis, Burkholderia pseudomallei and Brucella abortus. Thus, CLDC+MPF represents a novel prophylaxis to protect against multiple, highly virulent pathogens.
Conflict of interest statement
A provisional patent for CLDC+MPF has been filed by NIH on behalf of Catharine M. Bosio, John T. Belise and Jeff Fairman.
Figures
References
-
- Conlan JW, Chen W, Shen H, Webb A, KuoLee R. Experimental tularemia in mice challenged by aerosol or intradermally with virulent strains of Francisella tularensis: bacteriologic and histopathologic studies. Microb Pathog. 2003;34:239–248. - PubMed
-
- Saslaw S, Eigelsbach HT, Prior JA, Wilson HE, Carhart S. Tularemia vaccine study. II. Respiratory challenge. Arch Intern Med. 1961;107:702–714. - PubMed
-
- Cross JT, Jacobs RF. Tularemia: treatment failures with outpatient use of ceftriaxone. Clin Infect Dis. 1993;17:976–980. - PubMed
-
- Overholt EL, Tigertt WD, Kadull PJ, Ward MK, Charkes ND, et al. An analysis of forty-two cases of laboratory-acquired tularemia. Treatment with broad spectrum antibiotics. Am J Med. 1961;30:785–806. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
