

Review
doi: 10.1089/ars.2010.3241.
Epub 2010 Aug 30.
Nonhuman primate models of human immunology
Affiliations
- PMID: 20524846
- PMCID: PMC3014769
- DOI: 10.1089/ars.2010.3241
Item in Clipboard
Review
Nonhuman primate models of human immunology
Antioxid Redox Signal.
.
Abstract
Nonhuman primates have been used for biomedical research for several decades. The high level of genetic homology to humans coupled with their outbred nature has made nonhuman primates invaluable preclinical models. In this review, we summarize recent advances in our understanding of the nonhuman primate immune system, with special emphasis on studies carried out in rhesus macaque (Macaca mulatta). We highlight the utility of nonhuman primates in the characterization of immune senescence and the evaluation of new interventions to slow down the aging of the immune system.
Figures
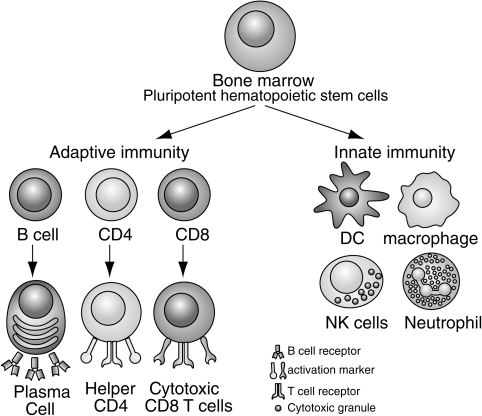
The immune system can be broadly divided into innate and adaptive branches. This figure highlights the key cellular components of each branch. Innate immunity is mostly mediated by dendritic cells (DC), natural killer (NK) cells, macrophages, and neutrophils. These cells use germline-encoded receptors to recognize pathogens. Adaptive immune response is mediated by T and B cells that express antigen receptors that recognize specific pathogens. The functions of these cells are discussed in more detail throughout the review.
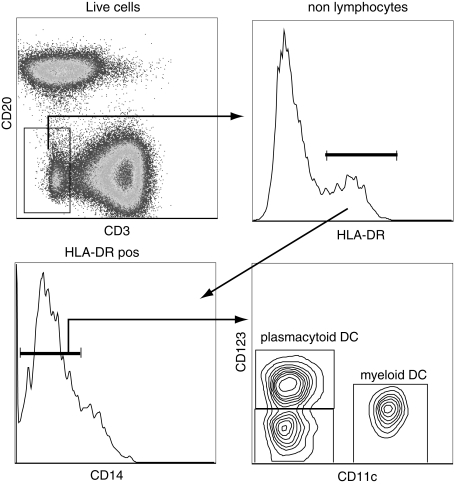
Identification of dendritic cell subsets in rhesus macaques. Dendritic cells (DCs) can be identified as nonlymphocytes (CD3negCD20neg) that express MHC-II molecules (HLA-DRpos) and lack the expression of CD14. DCs can be subdivided into myeloid DC and plasmacytoid DC based on the expression of CD11c and CD123. A minor subset does not express either of these markers.
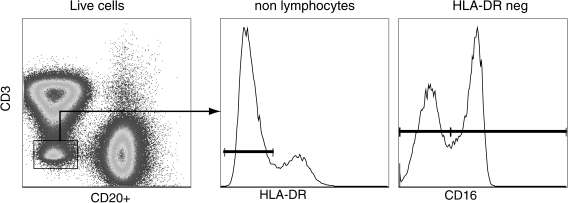
Identification of circulating natural killer cells in rhesus macaques. Peripheral blood resident NK cells can be identified as CD3negCD20negDRneg cells. The NK cells can then be subdivided into two subsets based on the expression of CD16: cytolytic NK cells (CD16pos); and inflammatory-cytokine producing NK cells (CD16neg).
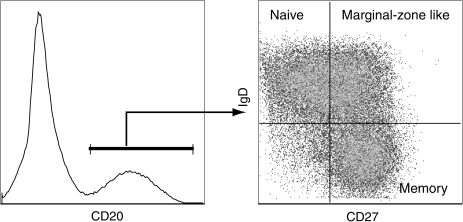
Delineation of B cell subsets in rhesus macaques. As described for humans, circulating B cells can be subdivided into three subsets based on the expression of CD27 and IgD. Naive B cells are identified as IgDposCD27neg and memory B cells are IgDnegCD27pos. In addition to these two main subsets, a third transitional subset referred as marginal zone-like B cells can be identified as IgDpos CD27pos. Both memory and marginal-zone like B cells contain somatic mutations in their B cell receptors, indicative of the fact that they have responded to antigens.
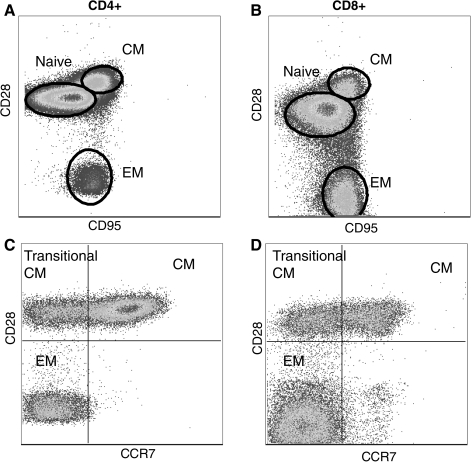
Delineation of rhesus macaque T cell subsets. T cells can be broadly subdivided into three subsets based on the expression of CD28 and CD95. Naive T cells are identified as CD28posCD95neg, central memory T cells are CD28posCD95neg, and effector memory T cell are CD28negCD95neg (A and B). The definition of memory T cells can be further refined based on the expression of CCR7 as described for human T cells. (C) and (D) show CD95pos CD4 and CD8 memory T cells. The expression of CCR7 together with CD28 allows the identification of central memory, effector memory, and transitional central memory T cells that are differentiating to effector memory.
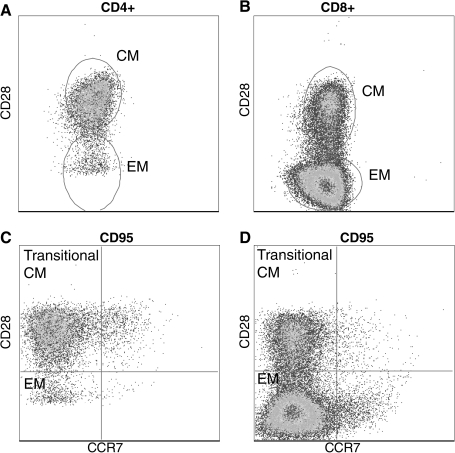
Nonlymphoid T cells do not express CCR7. T cells isolated from bronchial alveolar lavage are all memory phenotype based on CD95 expression. Furthermore, BAL-resident T cells lack the expression of CCR7.
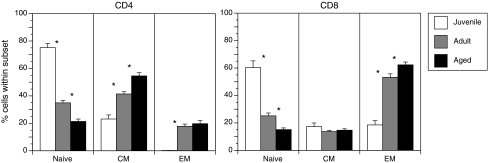
Advanced age is accompanied by severe loss of naive T cells in rhesus macaques. Frequency of naive, central memory (CM), and effector memory (EM) T cells was determined based on the expression of CD28 and CD95 in juvenile (∼1 year of age, n = 12), adult (5–10 years of age, n = 35) and aged (>17 years of age, n = 25). Cross sectional analysis indicates that, as described for humans, increasing age is associated with the loss of circulating naive T cells. Thus, rhesus macaques are a robust model to study mechanisms underlying T cell senescence.
Similar articles
-
Nonhuman primate models in the genomic era: a paradigm shift.ILAR J. 2013;54(2):154-65. doi: 10.1093/ilar/ilt044. ILAR J. 2013. PMID: 24174439 Free PMC article. Review.
-
Contributions of Nonhuman Primates to Research on Aging.Vet Pathol. 2016 Mar;53(2):277-90. doi: 10.1177/0300985815622974. Epub 2016 Feb 11. Vet Pathol. 2016. PMID: 26869153 Free PMC article. Review.
-
Immune senescence in aged nonhuman primates.Exp Gerontol. 2010 Sep;45(9):655-61. doi: 10.1016/j.exger.2010.06.001. Epub 2010 Jun 15. Exp Gerontol. 2010. PMID: 20558288 Free PMC article. Review.
-
Flow Cytometry-Based Methods to Characterize Immune Senescence in Nonhuman Primates.Methods Mol Biol. 2015;1343:65-80. doi: 10.1007/978-1-4939-2963-4_6. Methods Mol Biol. 2015. PMID: 26420709
-
Psychosocial factors and immune senescence in the aged primate.Ann N Y Acad Sci. 1992 Apr 15;650:276-82. doi: 10.1111/j.1749-6632.1992.tb49136.x. Ann N Y Acad Sci. 1992. PMID: 1605484
Cited by
-
The TLR2/TLR6 ligand FSL-1 mitigates radiation-induced hematopoietic injury in mice and nonhuman primates.Proc Natl Acad Sci U S A. 2023 Dec 12;120(50):e2122178120. doi: 10.1073/pnas.2122178120. Epub 2023 Dec 5. Proc Natl Acad Sci U S A. 2023. PMID: 38051771 Free PMC article.
-
Nonhuman Primates in Translational Research.Annu Rev Anim Biosci. 2022 Feb 15;10:441-468. doi: 10.1146/annurev-animal-021419-083813. Annu Rev Anim Biosci. 2022. PMID: 35167321 Free PMC article. Review.
-
In vivo characterization of alveolar and interstitial lung macrophages in rhesus macaques: implications for understanding lung disease in humans.J Immunol. 2014 Mar 15;192(6):2821-9. doi: 10.4049/jimmunol.1302269. Epub 2014 Feb 17. J Immunol. 2014. PMID: 24534529 Free PMC article.
-
Transplacental Antibody Transfer of Respiratory Syncytial Virus Specific IgG in Non-Human Primate Mother-Infant Pairs.Pathogens. 2021 Nov 5;10(11):1441. doi: 10.3390/pathogens10111441. Pathogens. 2021. PMID: 34832599 Free PMC article.
-
Social ageing can protect against infectious disease in a group-living primate.Philos Trans R Soc Lond B Biol Sci. 2024 Dec 16;379(1916):20220462. doi: 10.1098/rstb.2022.0462. Epub 2024 Oct 28. Philos Trans R Soc Lond B Biol Sci. 2024. PMID: 39463240 Free PMC article.
References
-
- Andrew D. Aspinall R. Il-7 and not stem cell factor reverses both the increase in apoptosis and the decline in thymopoiesis seen in aged mice. J Immunol. 2001;166:1524–1530. - PubMed
-
- Arstila TP. Casrouge A. Baron V. Even J. Kanellopoulos J. Kourilsky P. Diversity of human alpha beta T cell receptors. Science. 2000;288:1135. - PubMed
-
- Aspinall R. Mitchell W. Reversal of age-associated thymic atrophy: Treatments, delivery, and side effects. Exp Gerontol. 2008;43:700–705. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
