

Arabidopsis RETINOBLASTOMA-RELATED is required for stem cell maintenance, cell differentiation, and lateral organ production
- PMID: 20525851
- PMCID: PMC2910961
- DOI: 10.1105/tpc.110.074591
Arabidopsis RETINOBLASTOMA-RELATED is required for stem cell maintenance, cell differentiation, and lateral organ production
Abstract
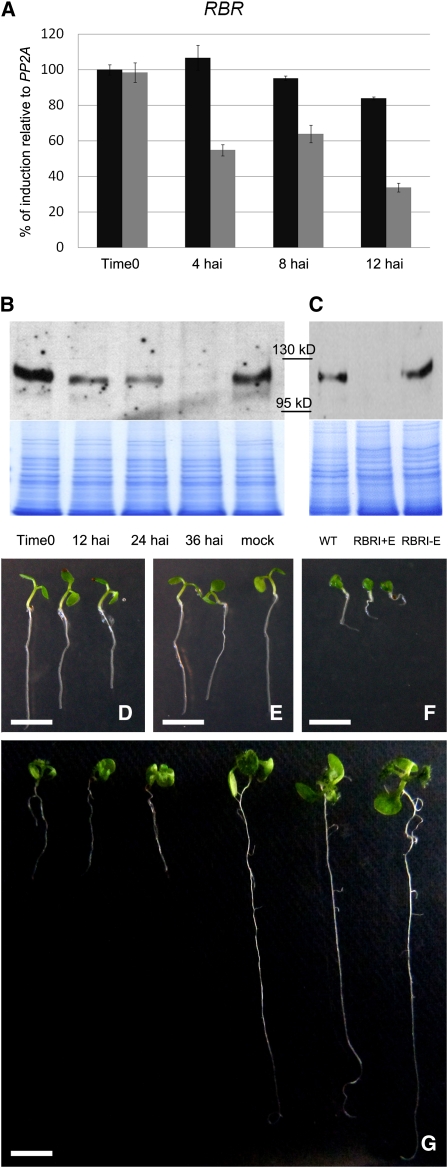
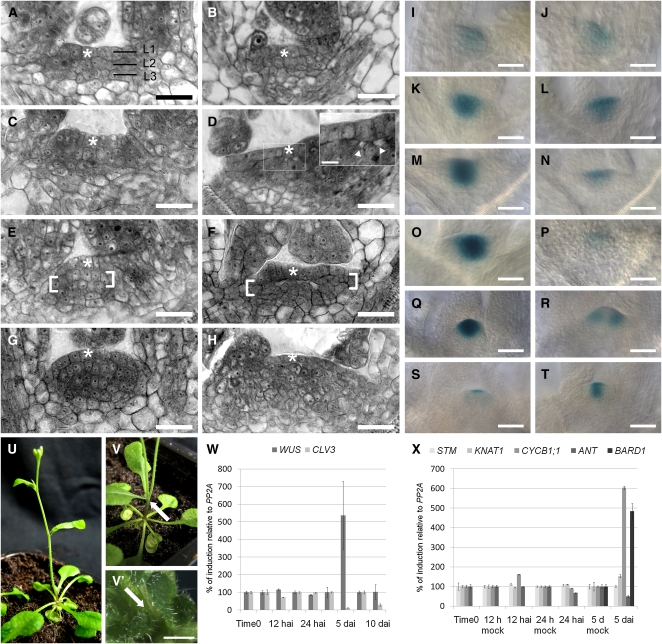
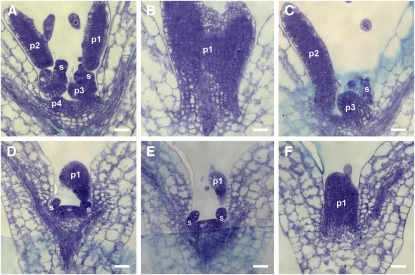
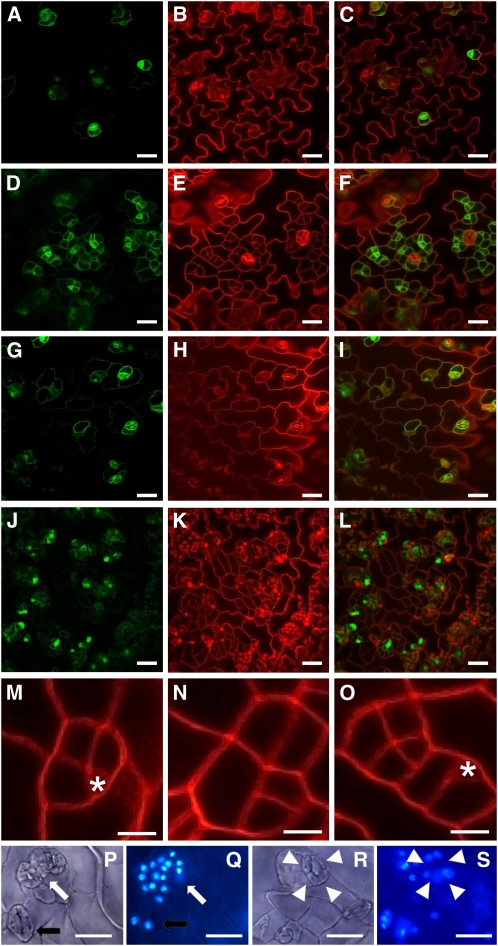
Several genes involved in the regulation of postembryonic organ initiation and growth have been identified. However, it remains largely unclear how developmental cues connect to the cell cycle. RETINOBLASTOMA RELATED (RBR) is a plant homolog of the tumor suppressor Retinoblastoma (pRb), which is a key regulator of the cell cycle. Using inducible RNA interference (RNAi) against Arabidopsis thaliana RBR (RBRi), we reduced RBR expression levels at different stages of plant development. Conditional reduction or loss of RBR function disrupted cell division patterns, promoted context-dependent cell proliferation, and negatively influenced establishment of cell differentiation. Several lineages of toti- and pluripotent cells, including shoot apical meristem stem cells, meristemoid mother cells, and procambial cells, failed to produce appropriately differentiated cells. Meristem activity was altered, leading to a disruption of the CLAVATA-WUSCHEL feedback loop and inhibition of lateral organ formation. Release of RBR from RNAi downregulation restored meristem activity. Gene profiling analyses soon after RBRi induction revealed that a change in RBR homeostasis is perceived as a stress, even before genes regulated by RBR-E2F become deregulated. The results establish RBR as a key cell cycle regulator required for coordination of cell division, differentiation, and cell homeostasis.
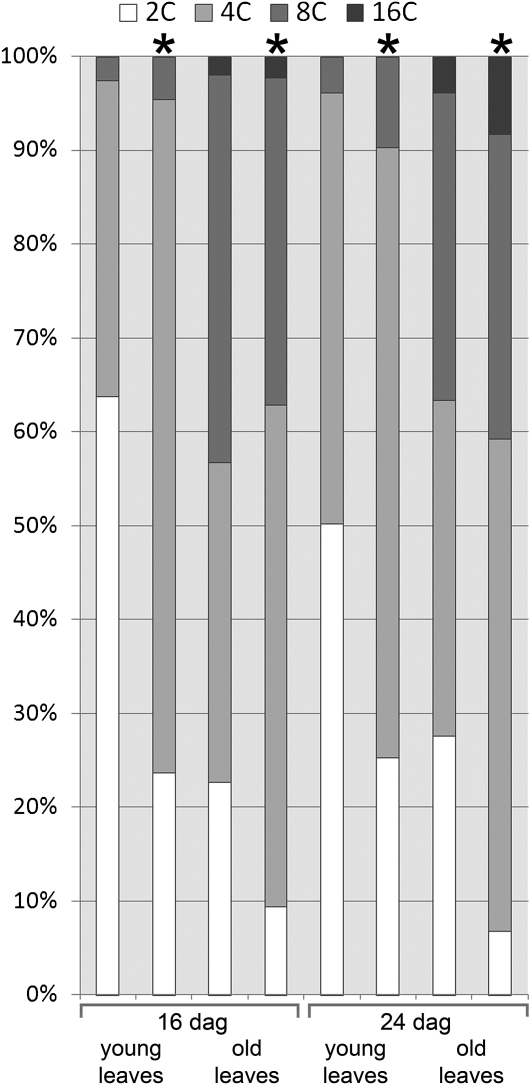
Figures



Similar articles
-
Induction of differentiation in the shoot apical meristem by transient overexpression of a retinoblastoma-related protein.Plant Physiol. 2006 Aug;141(4):1338-48. doi: 10.1104/pp.106.083022. Epub 2006 Jun 30. Plant Physiol. 2006. PMID: 16815954 Free PMC article.
-
Cell type-specific role of the retinoblastoma/E2F pathway during Arabidopsis leaf development.Plant Physiol. 2006 Jan;140(1):67-80. doi: 10.1104/pp.105.071027. Epub 2005 Dec 16. Plant Physiol. 2006. PMID: 16361519 Free PMC article.
-
Increased leaf mesophyll porosity following transient retinoblastoma-related protein silencing is revealed by microcomputed tomography imaging and leads to a system-level physiological response to the altered cell division pattern.Plant J. 2013 Dec;76(6):914-29. doi: 10.1111/tpj.12342. Epub 2013 Nov 11. Plant J. 2013. PMID: 24118480 Free PMC article.
-
Maintenance of stem cell populations in plants.Proc Natl Acad Sci U S A. 2003 Sep 30;100 Suppl 1(Suppl 1):11823-9. doi: 10.1073/pnas.1834206100. Epub 2003 Aug 20. Proc Natl Acad Sci U S A. 2003. PMID: 12930889 Free PMC article. Review.
-
Arabidopsis RETINOBLASTOMA-RELATED and Polycomb group proteins: cooperation during plant cell differentiation and development.J Exp Bot. 2014 Jun;65(10):2667-76. doi: 10.1093/jxb/eru069. Epub 2014 Mar 17. J Exp Bot. 2014. PMID: 24638900 Review.
Cited by
-
Low Temperature Affects Stem Cell Maintenance in Brassica oleracea Seedlings.Front Plant Sci. 2016 Jun 8;7:800. doi: 10.3389/fpls.2016.00800. eCollection 2016. Front Plant Sci. 2016. PMID: 27375654 Free PMC article.
-
Arabidopsis E2FA stimulates proliferation and endocycle separately through RBR-bound and RBR-free complexes.EMBO J. 2012 Mar 21;31(6):1480-93. doi: 10.1038/emboj.2012.13. Epub 2012 Feb 3. EMBO J. 2012. PMID: 22307083 Free PMC article.
-
Overexpression Populus d-Type Cyclin Gene PsnCYCD1;1 Influences Cell Division and Produces Curved Leaf in Arabidopsis thaliana.Int J Mol Sci. 2021 May 29;22(11):5837. doi: 10.3390/ijms22115837. Int J Mol Sci. 2021. PMID: 34072501 Free PMC article.
-
E2FB Interacts with RETINOBLASTOMA RELATED and Regulates Cell Proliferation during Leaf Development.Plant Physiol. 2020 Jan;182(1):518-533. doi: 10.1104/pp.19.00212. Epub 2019 Nov 6. Plant Physiol. 2020. PMID: 31694902 Free PMC article.
-
Plant tumors: a hundred years of study.Planta. 2020 Mar 18;251(4):82. doi: 10.1007/s00425-020-03375-5. Planta. 2020. PMID: 32189080 Review.
References
-
- Amirsadeghi S., McDonald A.E., Vanlerberghe G.C. (2007). A glucocorticoid-inducible gene expression system can cause growth defects in tobacco. Planta 226: 453–463 - PubMed
-
- Andersen S.U., Cvitanich C., Hougaard B.K., Roussis A., Gronlund M., Jensen D.B., Frokjaer L.A., Jensen E.O. (2003). The glucocorticoid-inducible GVG system causes severe growth defects in both root and shoot of the model legume Lotus japonicus. Mol. Plant Microbe Interact. 16: 1069–1076 - PubMed
-
- Aoyama T., Chua N.H. (1997). A glucocorticoid-mediated transcriptional induction system in transgenic plants. Plant J. 11: 605–612 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
