

Transmural heterogeneity of repolarization and Ca2+ handling in a model of mouse ventricular tissue
- PMID: 20525874
- PMCID: PMC2931428
- DOI: 10.1152/ajpheart.00907.2009
Transmural heterogeneity of repolarization and Ca2+ handling in a model of mouse ventricular tissue
Abstract
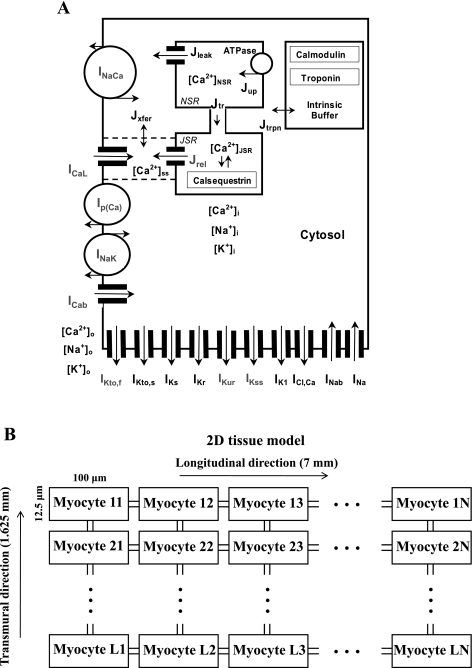
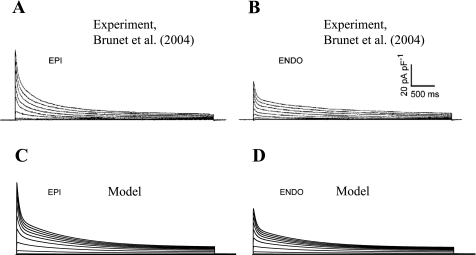
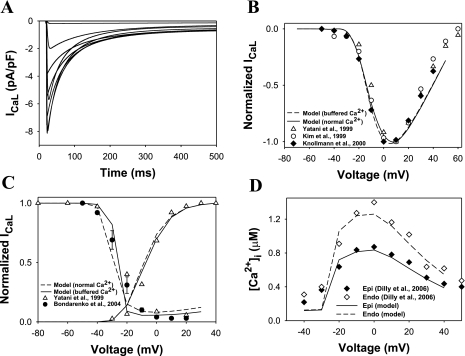
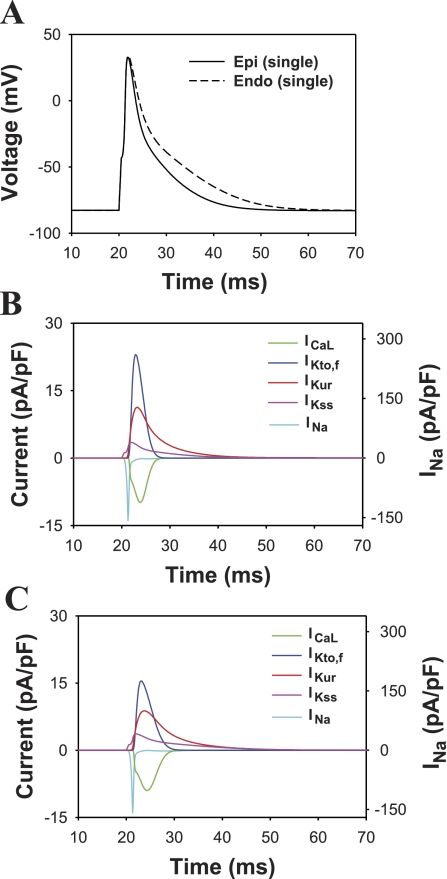
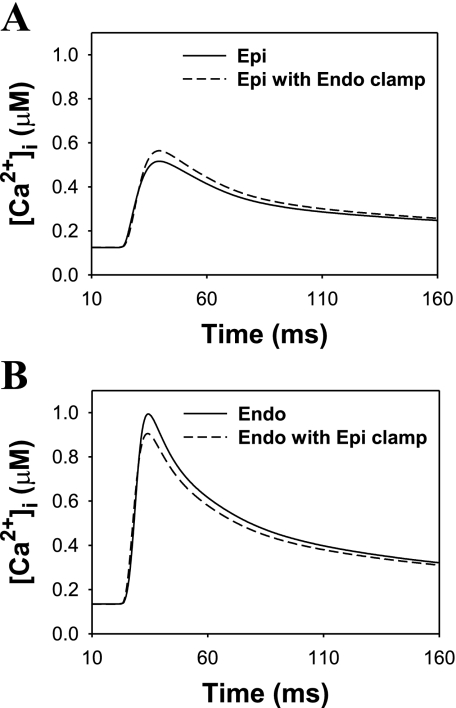
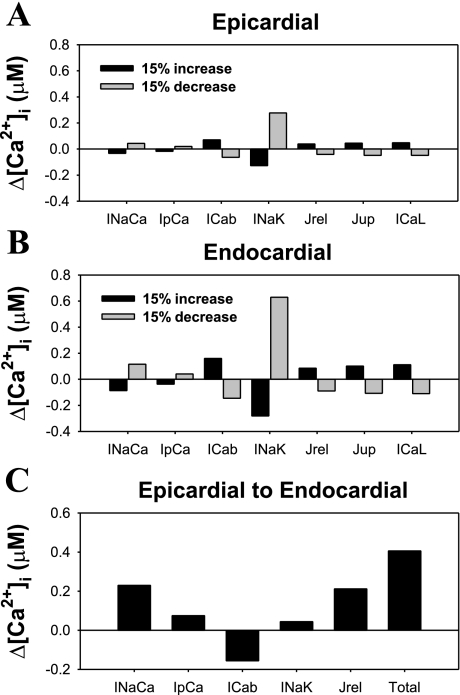
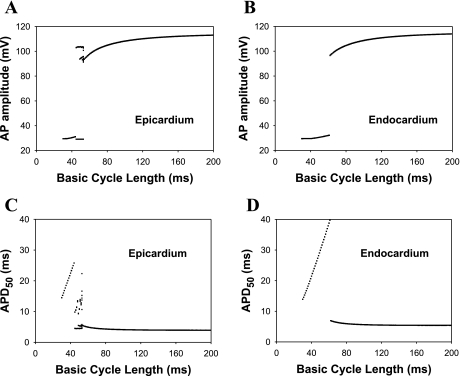
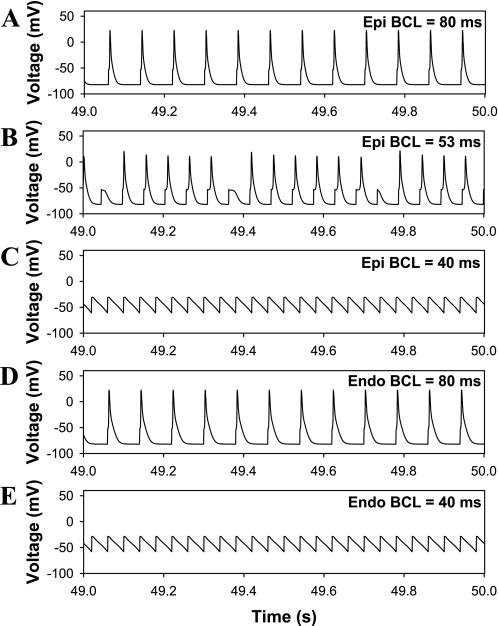
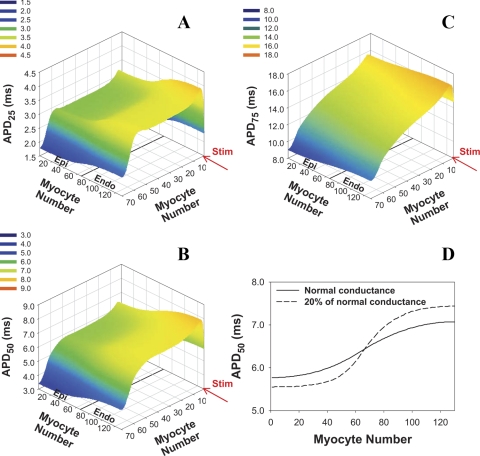
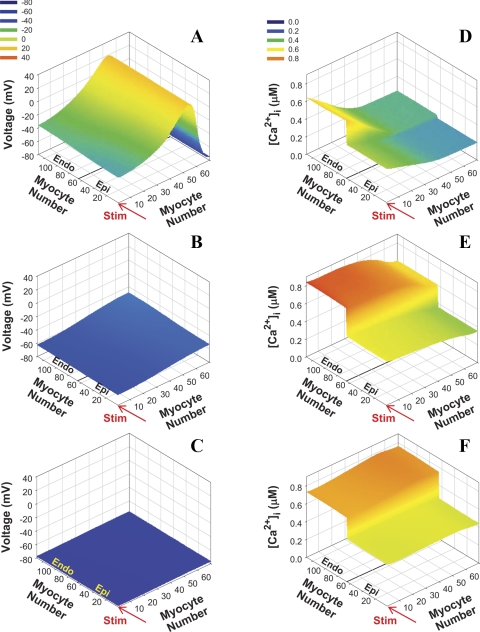
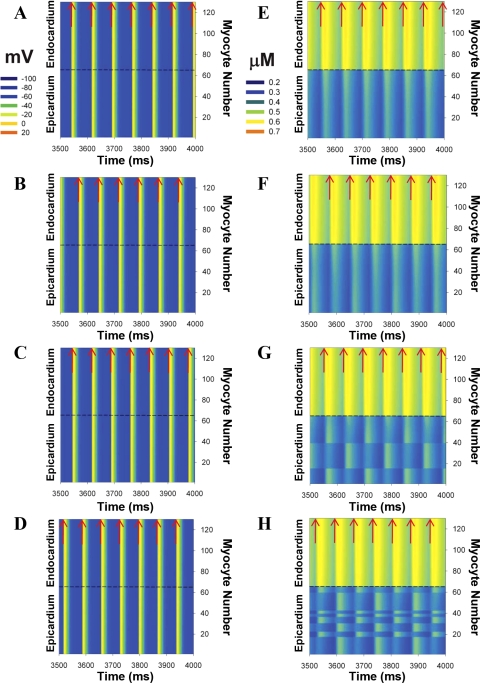
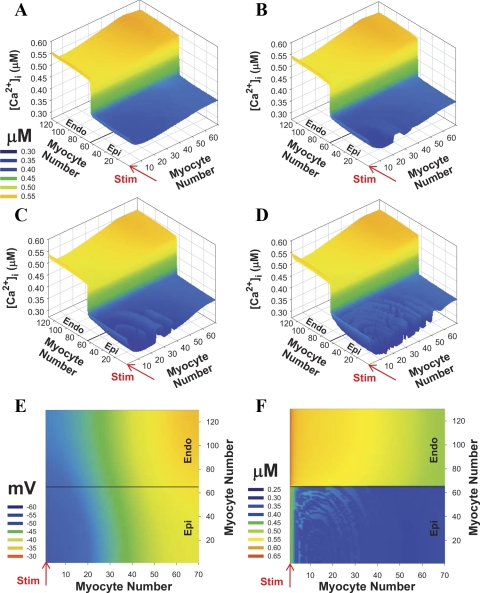
Mouse hearts have a diversity of action potentials (APs) generated by the cardiac myocytes from different regions. Recent evidence shows that cells from the epicardial and endocardial regions of the mouse ventricle have a diversity in Ca(2+) handling properties as well as K(+) current expression. To examine the mechanisms of AP generation, propagation, and stability in transmurally heterogeneous tissue, we developed a comprehensive model of the mouse cardiac cells from the epicardial and endocardial regions of the heart. Our computer model simulates the following differences between epicardial and endocardial myocytes: 1) AP duration is longer in endocardial and shorter in epicardial myocytes, 2) diastolic and systolic intracellular Ca(2+) concentration and intracellular Ca(2+) concentration transients are higher in paced endocardial and lower in epicardial myocytes, 3) Ca(2+) release rate is about two times larger in endocardial than in epicardial myocytes, and 4) Na(+)/Ca(2+) exchanger rate is greater in epicardial than in endocardial myocytes. Isolated epicardial cells showed a higher threshold for stability of AP generation but more complex patterns of AP duration at fast pacing rates. AP propagation velocities in the model of two-dimensional tissue are close to those measured experimentally. Simulations show that heterogeneity of repolarization and Ca(2+) handling are sustained across the mouse ventricular wall. Stability analysis of AP propagation in the two-dimensional model showed the generation of Ca(2+) alternans and more complex transmurally heterogeneous irregular structures of repolarization and intracellular Ca(2+) transients at fast pacing rates.
Figures
References
-
- Antzelevitch C, Shimizu W. Cellular mechanisms underlying the long QT syndrome. Curr Opin Cardiol 17: 43–51, 2002 - PubMed
-
- Anumonwo JM, Tallini YN, Vetter FJ, Jalife J. Action potential characteristics and arrhythmogenic properties of the cardiac conduction system of the murine heart. Circ Res 89: 329–335, 2001 - PubMed
-
- Bondarenko VE, Rasmusson RL. Re-entry in a multicellular model of mouse ventricle (Abstract). Biophys J 90: 547A, 2006
-
- Bondarenko VE, Rasmusson RL. Simulations of propagated mouse ventricular action potentials: effects of molecular heterogeneity. Am J Physiol Heart Circ Physiol 293: H1816–H1832, 2007 - PubMed
-
- Bondarenko VE, Rasmusson RL. Suppression of cellular alternans in guinea pig ventricular myocytes with LQT2: insights from the Luo-Rudy model. Int J Bifurcat Chaos 17: 381–425, 2007
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous