

Reliable semi-synthesis of hydrolysis-resistant 3'-peptidyl-tRNA conjugates containing genuine tRNA modifications
- PMID: 20525967
- PMCID: PMC2965236
- DOI: 10.1093/nar/gkq508
Reliable semi-synthesis of hydrolysis-resistant 3'-peptidyl-tRNA conjugates containing genuine tRNA modifications
Abstract
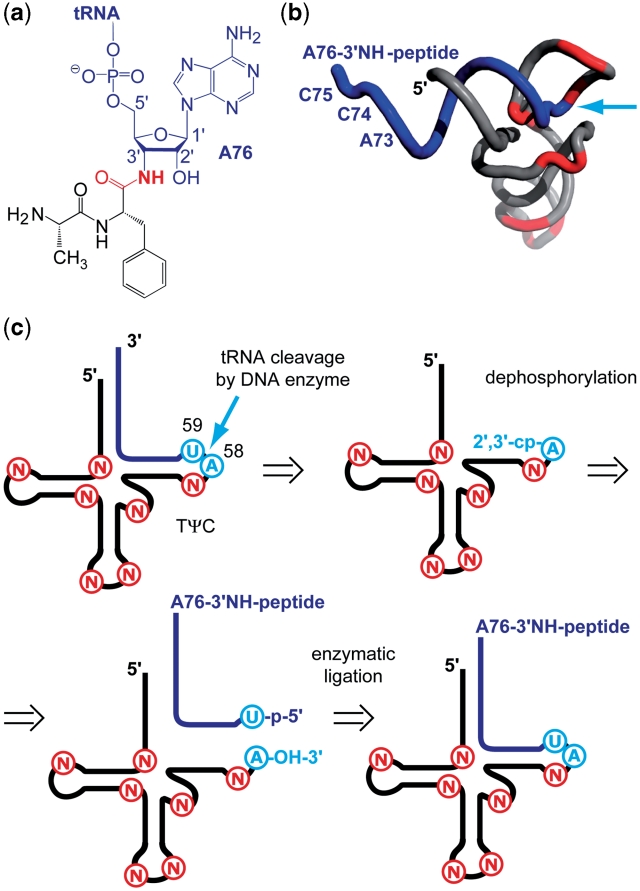
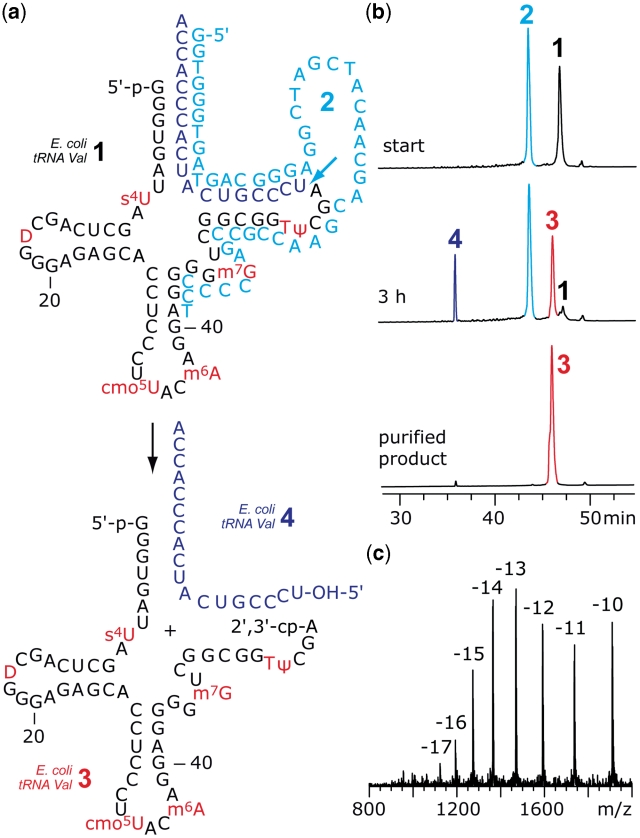
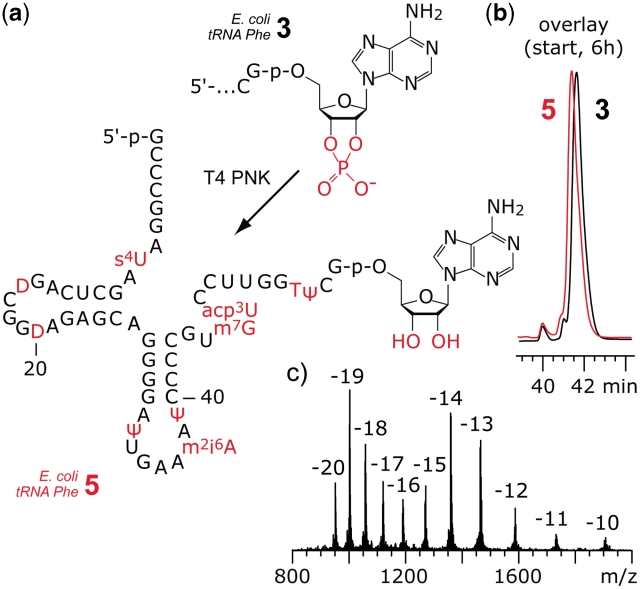
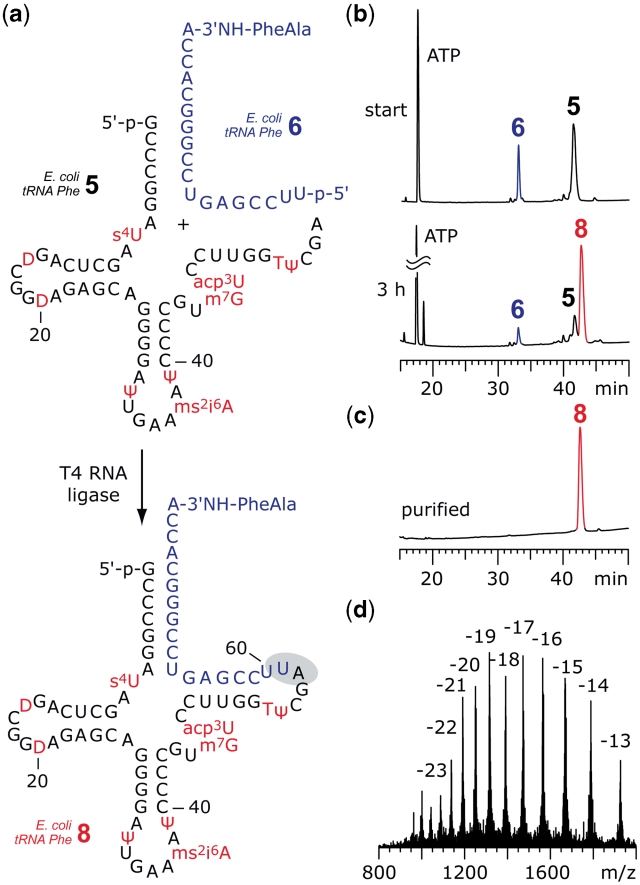
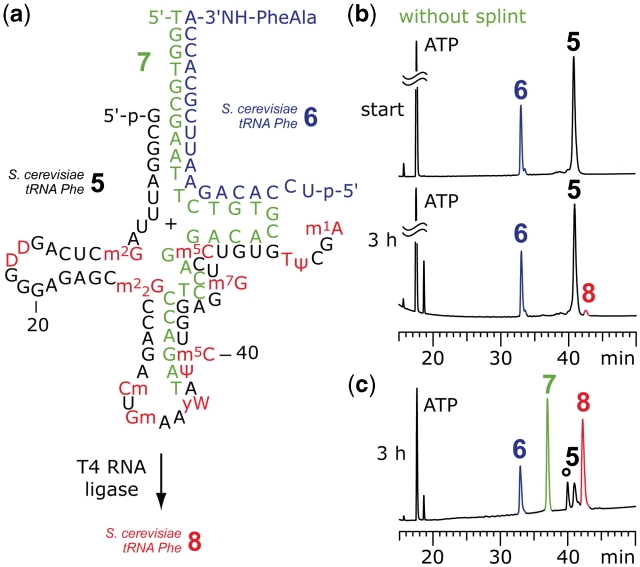
The 3'-peptidyl-tRNA conjugates that possess a hydrolysis-resistant ribose-3'-amide linkage instead of the natural ester linkage would represent valuable substrates for ribosomal studies. Up to date, access to these derivatives is severely limited. Here, we present a novel approach for the reliable synthesis of non-hydrolyzable 3'-peptidyl-tRNAs that contain all the respective genuine nucleoside modifications. In short, the approach is based on tRNAs from natural sources that are site-specifically cleaved within the TΨC loop by using DNA enzymes to obtain defined tRNA 5'-fragments carrying the modifications. After dephosphorylation of the 2',3'-cyclophosphate moieties from these fragments, they are ligated to the respective 3'-peptidylamino-tRNA termini that were prepared following the lines of a recently reported solid-phase synthesis. By this novel concept, non-hydrolyzable 3'-peptidyl-tRNA conjugates possessing all natural nucleoside modifications are accessible in highly efficient manner.
Figures
References
-
- Wilson DN, Nierhaus KH. The ribosome through the looking glass. Angew. Chem. Int. Ed. Engl. 2003;42:3464–3486. - PubMed
-
- Lang K, Erlacher M, Wilson DN, Micura R, Polacek N. The role of 23S ribosomal RNA residue A2451 in peptide bond synthesis revealed by atomic mutagenesis. Chem. Biol. 2008;15:485–492. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
