

Ciliary entry of the kinesin-2 motor KIF17 is regulated by importin-beta2 and RanGTP
- PMID: 20526328
- PMCID: PMC2896429
- DOI: 10.1038/ncb2073
Ciliary entry of the kinesin-2 motor KIF17 is regulated by importin-beta2 and RanGTP
Abstract
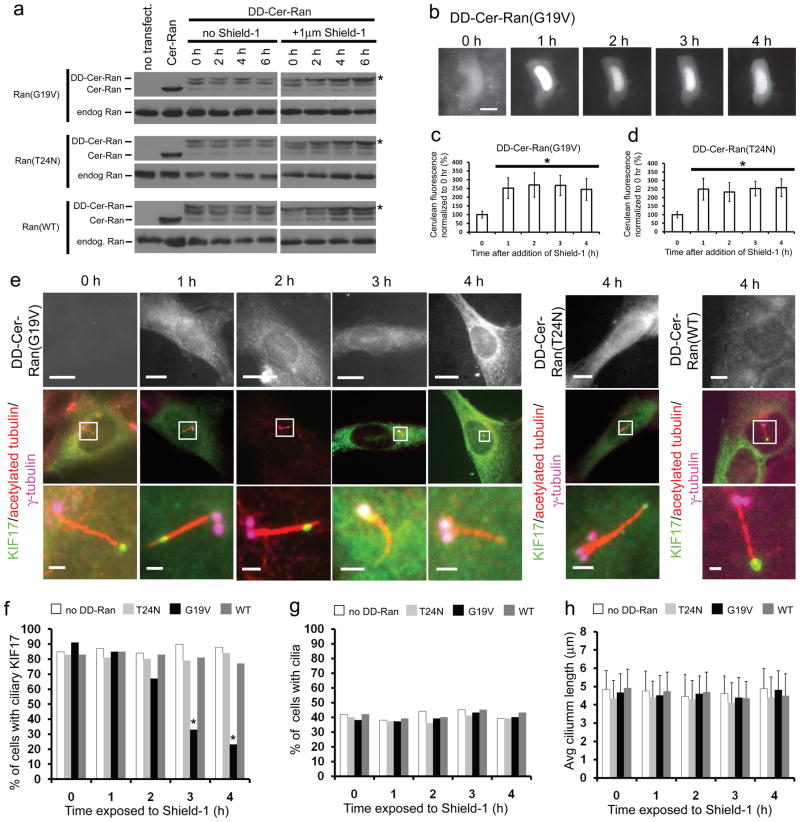
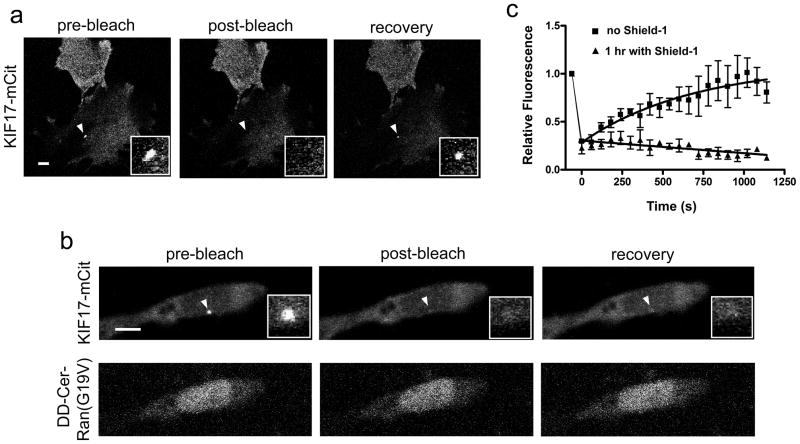
The biogenesis, maintenance and function of primary cilia are controlled through intraflagellar transport (IFT) driven by two kinesin-2 family members, the heterotrimeric KIF3A/KIF3B/KAP complex and the homodimeric KIF17 motor. How these motors and their cargoes gain access to the ciliary compartment is poorly understood. Here, we identify a ciliary localization signal (CLS) in the KIF17 tail domain that is necessary and sufficient for ciliary targeting. Similarities between the CLS and classic nuclear localization signals (NLSs) suggest that similar mechanisms regulate nuclear and ciliary import. We hypothesize that ciliary targeting of KIF17 is regulated by a ciliary-cytoplasmic gradient of the small GTPase Ran, with high levels of GTP-bound Ran (RanGTP) in the cilium. Consistent with this, cytoplasmic expression of GTP-locked Ran(G19V) disrupts the gradient and abolishes ciliary entry of KIF17. Furthermore, KIF17 interacts with the nuclear import protein importin-beta2 in a manner dependent on the CLS and inhibited by RanGTP. We propose that Ran has a global role in regulating cellular compartmentalization by controlling the shuttling of cytoplasmic proteins into nuclear and ciliary compartments.
Conflict of interest statement
The authors declare no competing financial interests.
Figures



Comment in
-
Nuclear transport receptor goes moonlighting.Nat Cell Biol. 2010 Jul;12(7):640-1. doi: 10.1038/ncb2076. Nat Cell Biol. 2010. PMID: 20526327
References
REFERENCES (MAIN TEXT)
-
- Silverman MA, Leroux MR. Intraflagellar transport and the generation of dynamic, structurally and functionally diverse cilia. Trends Cell Biol. 2009;19:306–316. - PubMed
-
- Satir P, Mitchell DR, Jekely G. How Did the Cilium Evolve? Ciliary Function in Mammalian Development. 2008;85:63–82. - PubMed
-
- Satir P, Christensen ST. Overview of structure and function of mammalian cilia. Annu Rev Physiol. 2007;69:377–400. - PubMed
REFERENCES (METHODS SECTION)
-
- Mayer U, et al. Proteomic analysis of a membrane preparation from rat olfactory sensory cilia. Chem Senses. 2008;33:145–162. - PubMed
-
- Mayer U, et al. The proteome of rat olfactory sensory cilia. Proteomics. 2009;9:322–334. - PubMed
-
- Davies S, Forge A. Preparation of the Mammalian Organ of Corti for Scanning Electron-Microscopy. J Microsc-Oxford. 1987;147:89–101. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- F32GM089034/GM/NIGMS NIH HHS/United States
- T32 GM007767/GM/NIGMS NIH HHS/United States
- R01GM070862/GM/NIGMS NIH HHS/United States
- R01 GM070862/GM/NIGMS NIH HHS/United States
- F31 DC009524/DC/NIDCD NIH HHS/United States
- R01GM083254/GM/NIGMS NIH HHS/United States
- T32 DC000011/DC/NIDCD NIH HHS/United States
- T32GM007767/GM/NIGMS NIH HHS/United States
- R01DC009606/DC/NIDCD NIH HHS/United States
- F32 GM089034/GM/NIGMS NIH HHS/United States
- F31DC009524/DC/NIDCD NIH HHS/United States
- R01 DC009606/DC/NIDCD NIH HHS/United States
- T32DC00011/DC/NIDCD NIH HHS/United States
- R01DK084725/DK/NIDDK NIH HHS/United States
- R01 DK084725/DK/NIDDK NIH HHS/United States
- R01 GM083254/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
