

Heme-heme and heme-ligand interactions in the di-heme oxygen-reducing site of cytochrome bd from Escherichia coli revealed by nanosecond absorption spectroscopy
- PMID: 20529691
- PMCID: PMC3990236
- DOI: 10.1016/j.bbabio.2010.05.010
Heme-heme and heme-ligand interactions in the di-heme oxygen-reducing site of cytochrome bd from Escherichia coli revealed by nanosecond absorption spectroscopy
Abstract
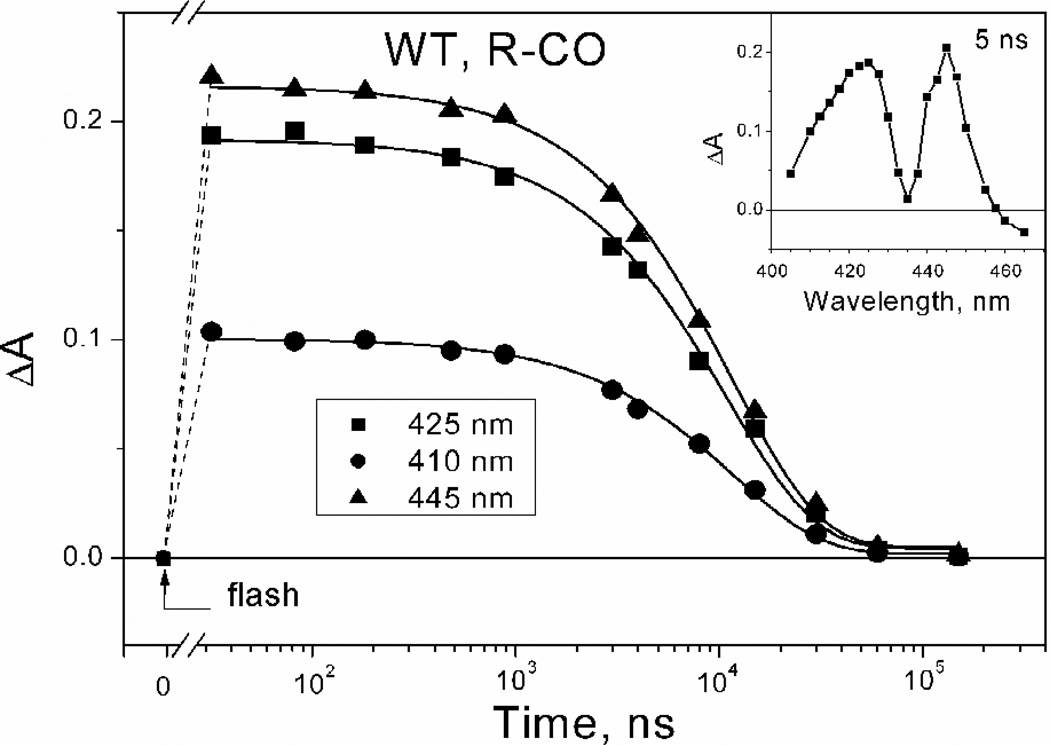
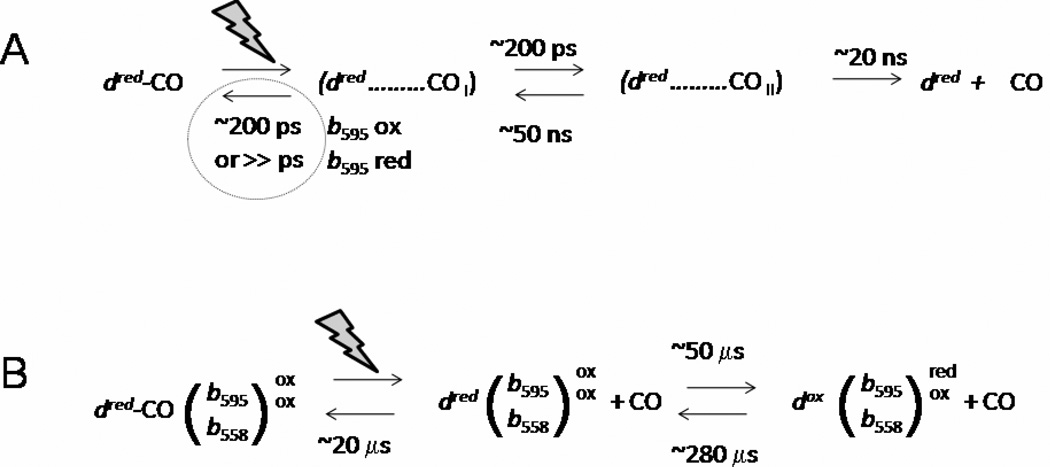
Cytochrome bd is a terminal quinol:O(2) oxidoreductase of respiratory chains of many bacteria. It contains three hemes, b(558), b(595), and d. The role of heme b(595) remains obscure. A CO photolysis/recombination study of the membranes of Escherichia coli containing either wild type cytochrome bd or inactive E445A mutant was performed using nanosecond absorption spectroscopy. We compared photoinduced changes of heme d-CO complex in one-electron-reduced, two-electron-reduced, and fully reduced states of cytochromes bd. The line shape of spectra of photodissociation of one-electron-reduced and two-electron-reduced enzymes is strikingly different from that of the fully reduced enzyme. The difference demonstrates that in the fully reduced enzyme photolysis of CO from heme d perturbs ferrous heme b(595) causing loss of an absorption band centered at 435 nm, thus supporting interactions between heme b(595) and heme d in the di-heme oxygen-reducing site, in agreement with previous works. Photolyzed CO recombines with the fully reduced enzyme monoexponentially with tau approximately 12 micros, whereas recombination of CO with one-electron-reduced cytochrome bd shows three kinetic phases, with tau approximately 14 ns, 14 micros, and 280 micros. The spectra of the absorption changes associated with these components are different in line shape. The 14 ns phase, absent in the fully reduced enzyme, reflects geminate recombination of CO with part of heme d. The 14-micros component reflects bimolecular recombination of CO with heme d and electron backflow from heme d to hemes b in approximately 4% of the enzyme population. The final, 280-micros component, reflects return of the electron from hemes b to heme d and bimolecular recombination of CO in that population. The fact that even in the two-electron-reduced enzyme, a nanosecond geminate recombination is observed, suggests that namely the redox state of heme b(595), and not that of heme b(558), controls the pathway(s) by which CO migrates between heme d and the medium.
2010 Elsevier B.V. All rights reserved.
Figures


Similar articles
-
Microsecond time-resolved absorption spectroscopy used to study CO compounds of cytochrome bd from Escherichia coli.PLoS One. 2014 Apr 22;9(4):e95617. doi: 10.1371/journal.pone.0095617. eCollection 2014. PLoS One. 2014. PMID: 24755641 Free PMC article.
-
Femtosecond resolution of ligand-heme interactions in the high-affinity quinol oxidase bd: A di-heme active site?Proc Natl Acad Sci U S A. 2000 Feb 15;97(4):1554-9. doi: 10.1073/pnas.030528197. Proc Natl Acad Sci U S A. 2000. PMID: 10660685 Free PMC article.
-
Accommodation of CO in the di-heme active site of cytochrome bd terminal oxidase from Escherichia coli.J Inorg Biochem. 2013 Jan;118:65-7. doi: 10.1016/j.jinorgbio.2012.09.016. Epub 2012 Sep 24. J Inorg Biochem. 2013. PMID: 23123340
-
Oxygen as Acceptor.EcoSal Plus. 2015;6(2):10.1128/ecosalplus.ESP-0012-2015. doi: 10.1128/ecosalplus.ESP-0012-2015. EcoSal Plus. 2015. PMID: 26734697 Free PMC article. Review.
-
Probing molecular structure of dioxygen reduction site of bacterial quinol oxidases through ligand binding to the redox metal centers.J Inorg Biochem. 2000 Nov;82(1-4):19-25. doi: 10.1016/s0162-0134(00)00140-9. J Inorg Biochem. 2000. PMID: 11132627 Review.
Cited by
-
Aerobic respiratory chain of Escherichia coli is not allowed to work in fully uncoupled mode.Proc Natl Acad Sci U S A. 2011 Oct 18;108(42):17320-4. doi: 10.1073/pnas.1108217108. Epub 2011 Oct 10. Proc Natl Acad Sci U S A. 2011. PMID: 21987791 Free PMC article.
-
An Engineered Glutamate in Biosynthetic Models of Heme-Copper Oxidases Drives Complete Product Selectivity by Tuning the Hydrogen-Bonding Network.Biochemistry. 2021 Feb 2;60(4):346-355. doi: 10.1021/acs.biochem.0c00852. Epub 2021 Jan 19. Biochemistry. 2021. PMID: 33464878 Free PMC article.
-
Oxoferryl-porphyrin radical catalytic intermediate in cytochrome bd oxidases protects cells from formation of reactive oxygen species.J Biol Chem. 2012 Mar 16;287(12):8830-8. doi: 10.1074/jbc.M111.333542. Epub 2012 Jan 27. J Biol Chem. 2012. PMID: 22287551 Free PMC article.
-
Bacterial Oxidases of the Cytochrome bd Family: Redox Enzymes of Unique Structure, Function, and Utility As Drug Targets.Antioxid Redox Signal. 2021 Jun 1;34(16):1280-1318. doi: 10.1089/ars.2020.8039. Epub 2020 Nov 9. Antioxid Redox Signal. 2021. PMID: 32924537 Free PMC article. Review.
-
Cytochrome bd Displays Significant Quinol Peroxidase Activity.Sci Rep. 2016 Jun 9;6:27631. doi: 10.1038/srep27631. Sci Rep. 2016. PMID: 27279363 Free PMC article.
References
-
- Poole RK, Cook GM. Redundancy of aerobic respiratory chains in bacteria? Routes, reasons and regulation. Adv. Microb. Physiol. 2000;43:165–224. - PubMed
-
- Junemann S. Cytochrome bd terminal oxidase. Biochim. Biophys. Acta. 1997;1321:107–127. - PubMed
-
- Borisov VB. Cytochrome bd: structure and properties. Biochemistry (Moscow) 1996;61:565–574. (translated from Biokhimiya (in Russian) (1996), 61, 786–799). - PubMed
-
- Tsubaki M, Hori H, Mogi T. Probing molecular structure of dioxygen reduction site of bacterial quinol oxidases through ligand binding to the redox metal centers. J. Inorg. Biochem. 2000;82:19–25. - PubMed
-
- Borisov VB, Verkhovsky MI. Oxygen as acceptor [Chapter 3.2.7] In: Böck A, Curtiss R III, Kaper JB, Neidhardt FC, Nyström T, Rudd KE, Squires CL, editors. EcoSal - Escherichia coli and Salmonella: cellular and molecular biology. Washington, DC: ASM Press; 2009. < http://www.ecosal.org>.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
