

Mitochondria-targeted cytochrome P450 2E1 induces oxidative damage and augments alcohol-mediated oxidative stress
- PMID: 20529841
- PMCID: PMC2915697
- DOI: 10.1074/jbc.M110.121822
Mitochondria-targeted cytochrome P450 2E1 induces oxidative damage and augments alcohol-mediated oxidative stress
Abstract
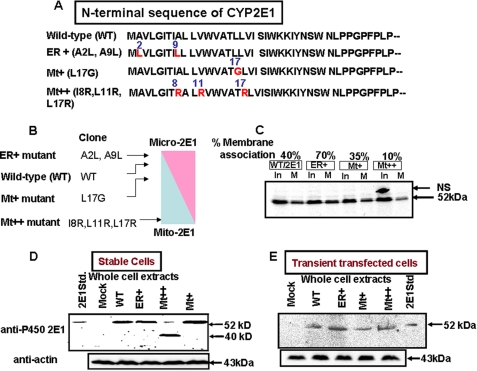
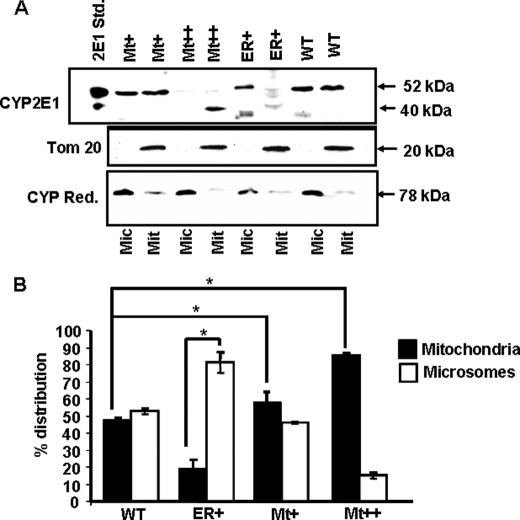
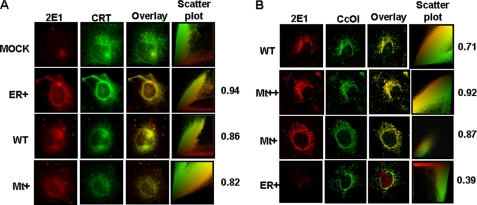
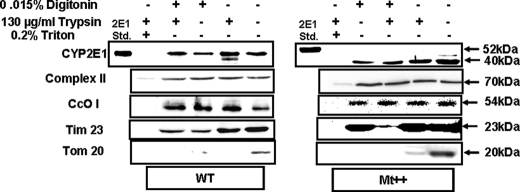
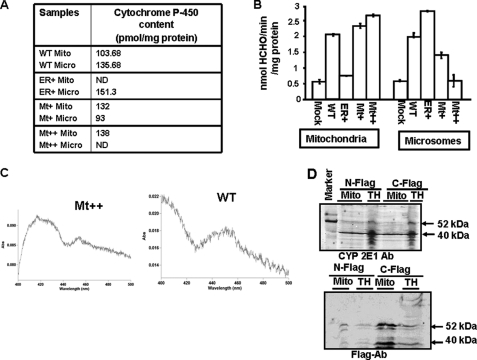
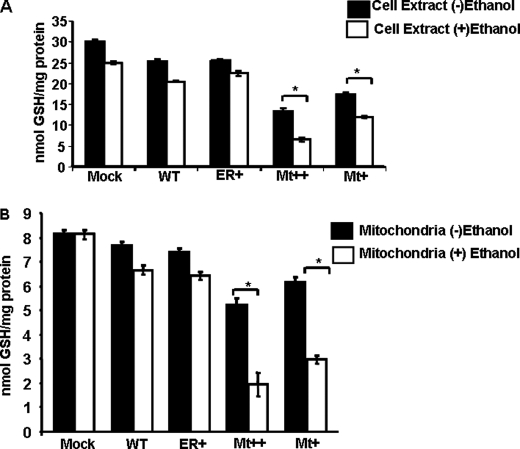
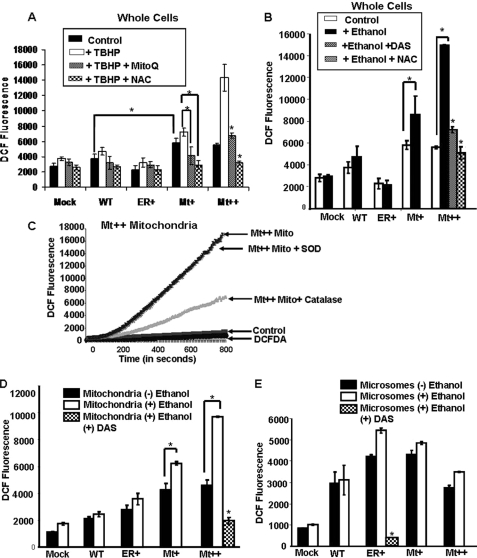
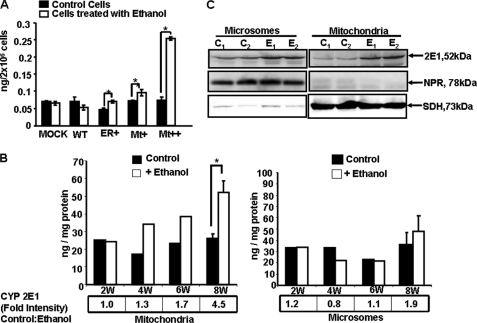
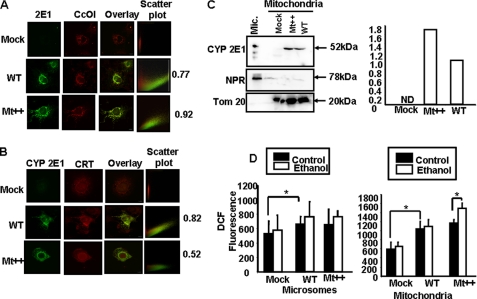
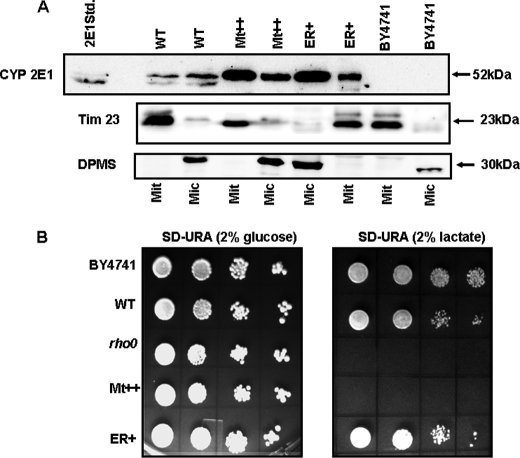
The ethanol-inducible cytochrome P450 2E1 (CYP2E1) is also induced under different pathological and physiological conditions. Studies including ours have shown that CYP2E1 is bimodally targeted to both the endoplasmic reticulum (microsomes) (mc CYP2E1) and mitochondria (mt CYP2E1). In this study we investigated the role of mtCYP2E1 in ethanol-mediated oxidative stress in stable cell lines expressing predominantly mt CYP2E1 or mc CYP2E1. The ER+ mutation (A2L, A9L), which increases the affinity of the nascent protein for binding to the signal recognition particle, preferentially targets CYP2E1 to the endoplasmic reticulum. The Mt+ (L17G) and Mt++ (I8R, L11R, L17R) mutant proteins, showing progressively lower affinity for signal recognition particle binding, were targeted to mitochondria at correspondingly higher levels. The rate of GSH depletion, used as a measure of oxidative stress, was higher in cells expressing Mt++ and Mt+ proteins as compared with cells expressing ER+ protein. In addition, the cellular level of F(2)-isoprostanes, a direct indicator of oxidative stress, was increased markedly in Mt++ cells after ethanol treatment. Notably, expression of Mt++ CYP2E1 protein in yeast cells caused more severe mitochondrial DNA damage and respiratory deficiency than the wild type or ER+ proteins as tested by the inability of cells to grow on glycerol or ethanol. Additionally, liver mitochondria from ethanol-fed rats containing high mt CYP2E1 showed higher levels of F(2)-isoprostane production. These results strongly suggest that mt CYP2E1 induces oxidative stress and augments alcohol-mediated cell/tissue injury.
Figures
Similar articles
-
Human cytochrome P450 2E1 mutations that alter mitochondrial targeting efficiency and susceptibility to ethanol-induced toxicity in cellular models.J Biol Chem. 2013 May 3;288(18):12627-44. doi: 10.1074/jbc.M113.452367. Epub 2013 Mar 7. J Biol Chem. 2013. PMID: 23471973 Free PMC article.
-
Role of CYP2E1 in Mitochondrial Dysfunction and Hepatic Injury by Alcohol and Non-Alcoholic Substances.Curr Mol Pharmacol. 2017;10(3):207-225. doi: 10.2174/1874467208666150817111114. Curr Mol Pharmacol. 2017. PMID: 26278393 Free PMC article. Review.
-
Additive effects of mitochondrion-targeted cytochrome CYP2E1 and alcohol toxicity on cytochrome c oxidase function and stability of respirosome complexes.J Biol Chem. 2012 May 4;287(19):15284-97. doi: 10.1074/jbc.M111.314062. Epub 2012 Mar 6. J Biol Chem. 2012. PMID: 22396533 Free PMC article.
-
Overexpression of CYP2E1 in mitochondria sensitizes HepG2 cells to the toxicity caused by depletion of glutathione.J Biol Chem. 2006 Feb 24;281(8):5128-36. doi: 10.1074/jbc.M510484200. Epub 2005 Dec 27. J Biol Chem. 2006. PMID: 16380384
-
Roles of Cytochrome P450 in Metabolism of Ethanol and Carcinogens.Adv Exp Med Biol. 2018;1032:15-35. doi: 10.1007/978-3-319-98788-0_2. Adv Exp Med Biol. 2018. PMID: 30362088 Free PMC article. Review.
Cited by
-
Treatment with omega-3 PUFAs does not increase the risk of CYP2E1-dependent oxidative stress and diabetic liver pathology.Front Endocrinol (Lausanne). 2022 Sep 26;13:1004564. doi: 10.3389/fendo.2022.1004564. eCollection 2022. Front Endocrinol (Lausanne). 2022. PMID: 36225205 Free PMC article.
-
Human cytochrome P450 2E1 mutations that alter mitochondrial targeting efficiency and susceptibility to ethanol-induced toxicity in cellular models.J Biol Chem. 2013 May 3;288(18):12627-44. doi: 10.1074/jbc.M113.452367. Epub 2013 Mar 7. J Biol Chem. 2013. PMID: 23471973 Free PMC article.
-
Mechanisms of drug toxicity and relevance to pharmaceutical development.Drug Metab Pharmacokinet. 2011;26(1):3-14. doi: 10.2133/dmpk.dmpk-10-rv-062. Epub 2010 Oct 22. Drug Metab Pharmacokinet. 2011. PMID: 20978361 Free PMC article. Review.
-
myo-Inositol Oxygenase Overexpression Accentuates Generation of Reactive Oxygen Species and Exacerbates Cellular Injury following High Glucose Ambience: A NEW MECHANISM RELEVANT TO THE PATHOGENESIS OF DIABETIC NEPHROPATHY.J Biol Chem. 2016 Mar 11;291(11):5688-5707. doi: 10.1074/jbc.M115.669952. Epub 2016 Jan 20. J Biol Chem. 2016. Retraction in: J Biol Chem. 2019 Jun 28;294(26):10380. doi: 10.1074/jbc.W119.009586. PMID: 26792859 Free PMC article. Retracted.
-
Role of CYP2E1 in Mitochondrial Dysfunction and Hepatic Injury by Alcohol and Non-Alcoholic Substances.Curr Mol Pharmacol. 2017;10(3):207-225. doi: 10.2174/1874467208666150817111114. Curr Mol Pharmacol. 2017. PMID: 26278393 Free PMC article. Review.
References
-
- Bondoc F. Y., Bao Z., Hu W. Y., Gonzalez F. J., Wang Y., Yang C. S., Hong J. Y. (1999) Biochem. Pharmacol. 58, 461–463 - PubMed
-
- Ronis M. J., Lindros K. O., Ingelman-Sundberg M. (1996) in Cytochromes P450, Pharmacological and Toxicological Aspects (Ioannides C. ed.) CRC Press. Inc., Boca Raton, FL
-
- Sohn O. S., Fiala E. S., Requeijo S. P., Weisburger J. H., Gonzalez F. J. (2001) Cancer Res. 61, 8435–8440 - PubMed
-
- Sumner S. C., Fennell T. R., Moore T. A., Chanas B., Gonzalez F., Ghanayem B. I. (1999) Chem. Res. Toxicol. 12, 1110–1116 - PubMed
-
- Zaher H., Buters J. T., Ward J. M., Bruno M. K., Lucas A. M., Stern S. T., Cohen S. D., Gonzalez F. J. (1998) Toxicol. Appl. Pharmacol. 152, 193–199 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
