

A chemotactic gradient sequestered on endothelial heparan sulfate induces directional intraluminal crawling of neutrophils
- PMID: 20530797
- PMCID: PMC3173988
- DOI: 10.1182/blood-2010-01-266072
A chemotactic gradient sequestered on endothelial heparan sulfate induces directional intraluminal crawling of neutrophils
Abstract
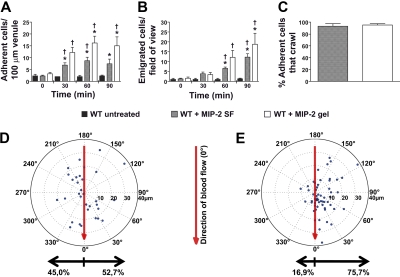
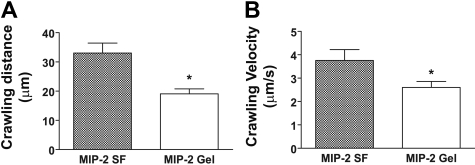
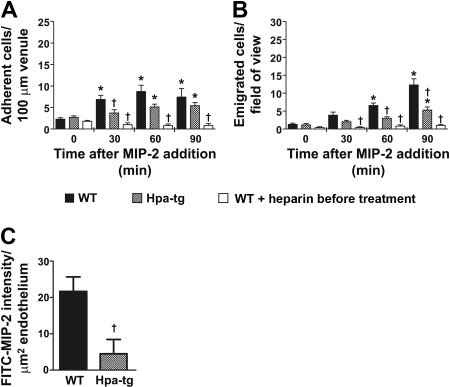
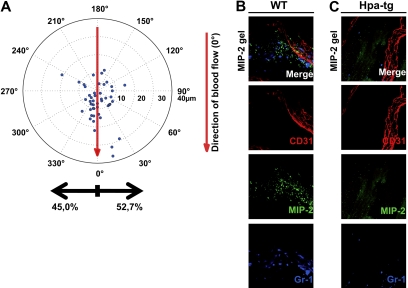
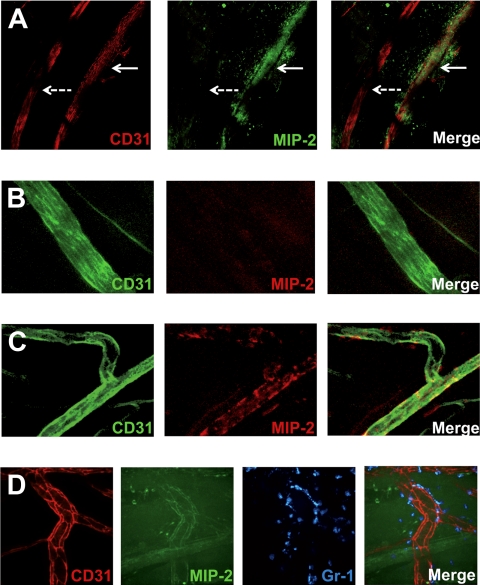
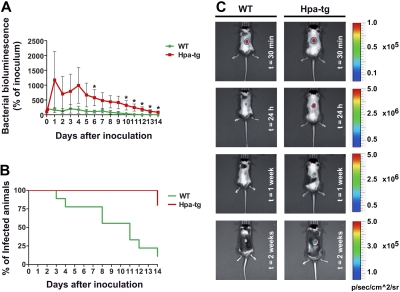
During infection, chemokines sequestered on endothelium induce recruitment of circulating leukocytes into the tissue where they chemotax along chemokine gradients toward the afflicted site. The aim of this in vivo study was to determine whether a chemokine gradient was formed intravascularly and influenced intraluminal neutrophil crawling and transmigration. A chemokine gradient was induced by placing a macrophage inflammatory protein-2 (MIP-2)-containing (CXCL2) gel on the cremaster muscle of anesthetized wild-type mice or heparanase-overexpressing transgenic mice (hpa-tg) with truncated heparan sulfate (HS) side chains. Neutrophil-endothelial interactions were visualized by intravital microscopy and chemokine gradients detected by confocal microscopy. Localized extravascular chemokine release (MIP-2 gel) induced directed neutrophil crawling along a chemotactic gradient immobilized on the endothelium and accelerated their recruitment into the target tissue compared with homogeneous extravascular chemokine concentration (MIP-2 superfusion). Endothelial chemokine sequestration occurred exclusively in venules and was HS-dependent, and neutrophils in hpa-tg mice exhibited random crawling. Despite similar numbers of adherent neutrophils in hpa-tg and wild-type mice, the altered crawling in hpa-tg mice was translated into decreased number of emigrated neutrophils and ultimately decreased the ability to clear bacterial infections. In conclusion, an intravascular chemokine gradient sequestered by endothelial HS effectively directs crawling leukocytes toward transmigration loci close to the infection site.
Figures
References
-
- Ley K, Laudanna C, Cybulsky MI, Nourshargh S. Getting to the site of inflammation: the leukocyte adhesion cascade updated. Nat Rev Immunol. 2007;7(9):678–689. - PubMed
-
- Petri B, Phillipson M, Kubes P. The physiology of leukocyte recruitment: an in vivo perspective. J Immunol. 2008;180(10):6439–6446. - PubMed
-
- Springer TA. Traffic signals for lymphocyte recirculation and leukocyte emigration: the multistep paradigm. Cell. 1994;76(2):301–314. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
