

Purinergic signaling: a fundamental mechanism in neutrophil activation
- PMID: 20530802
- PMCID: PMC4209711
- DOI: 10.1126/scisignal.2000549
Purinergic signaling: a fundamental mechanism in neutrophil activation
Abstract
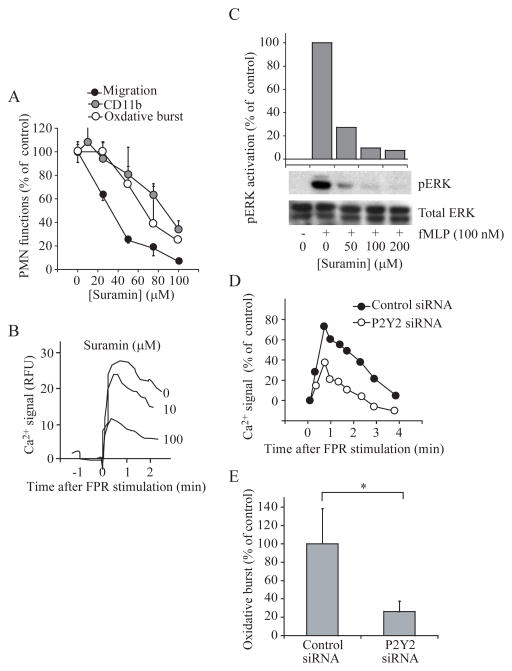
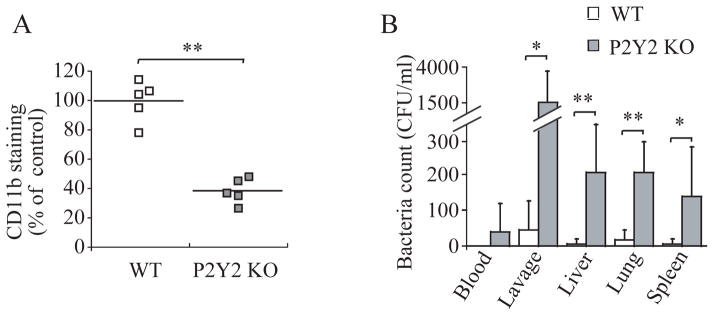
Efficient activation of neutrophils is a key requirement for effective immune responses. We found that neutrophils released cellular adenosine triphosphate (ATP) in response to exogenous stimuli such as formylated bacterial peptides and inflammatory mediators that activated Fcgamma, interleukin-8, C5a complement, and leukotriene B(4) receptors. Stimulation of the formyl peptide receptor (FPR) led to ATP release through pannexin-1 (panx1) hemichannels, and FPRs colocalized with P2Y2 nucleotide receptors on the cell surface to form a purinergic signaling system that facilitated neutrophil activation. Disruption of this purinergic signaling system by inhibiting or silencing panx1 hemichannels or P2Y2 receptors blocked neutrophil activation and impaired innate host responses to bacterial infection. Thus, purinergic signaling is a fundamental mechanism required for neutrophil activation and immune defense.
Conflict of interest statement
Figures





References
-
- Nussler AK, Wittel UA, Nussler NC, Beger HG. Leukocytes, the Janus cells in inflammatory disease. Langenbecks Arch Surg. 1999;384:222–232. - PubMed
-
- Waugh J, Wilson C. The interleukin-8 pathway in cancer. Clin Cancer Res. 2008;14:6735–6741. - PubMed
-
- Capra V. Molecular and functional aspects of human cysteinyl leukotriene receptors. Pharmacol Res. 2004;50:1–11. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
