

Hepcidin mediates transcriptional changes that modulate acute cytokine-induced inflammatory responses in mice
- PMID: 20530874
- PMCID: PMC2898601
- DOI: 10.1172/JCI42011
Hepcidin mediates transcriptional changes that modulate acute cytokine-induced inflammatory responses in mice
Abstract
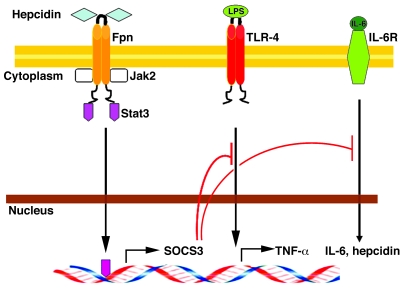
Hepcidin is a peptide hormone that regulates iron homeostasis and acts as an antimicrobial peptide. It is expressed and secreted by a variety of cell types in response to iron loading and inflammation. Hepcidin mediates iron homeostasis by binding to the iron exporter ferroportin, inducing its internalization and degradation via activation of the protein kinase Jak2 and the subsequent phosphorylation of ferroportin. Here we have shown that hepcidin-activated Jak2 also phosphorylates the transcription factor Stat3, resulting in a transcriptional response. Hepcidin treatment of ferroportin-expressing mouse macrophages showed changes in mRNA expression levels of a wide variety of genes. The changes in transcript levels for half of these genes were a direct effect of hepcidin, as shown by cycloheximide insensitivity, and dependent on the presence of Stat3. Hepcidin-mediated transcriptional changes modulated LPS-induced transcription in both cultured macrophages and in vivo mouse models, as demonstrated by suppression of IL-6 and TNF-alpha transcript and secreted protein. Hepcidin-mediated transcription in mice also suppressed toxicity and morbidity due to single doses of LPS, poly(I:C), and turpentine, which is used to model chronic inflammatory disease. Most notably, we demonstrated that hepcidin pretreatment protected mice from a lethal dose of LPS and that hepcidin-knockout mice could be rescued from LPS toxicity by injection of hepcidin. The results of our study suggest a new function for hepcidin in modulating acute inflammatory responses.
Figures





Comment in
-
The hepcidin circuits act: balancing iron and inflammation.Hepatology. 2011 May;53(5):1764-6. doi: 10.1002/hep.24267. Hepatology. 2011. PMID: 21520181 Free PMC article.
References
-
- Shi J, Camus AC. Hepcidins in amphibians and fishes: Antimicrobial peptides or iron-regulatory hormones? Dev Comp Immunol. 2006;30(9):746–755. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
