

Meiosis requires a translational positive loop where CPEB1 ensues its replacement by CPEB4
- PMID: 20531391
- PMCID: PMC2905248
- DOI: 10.1038/emboj.2010.111
Meiosis requires a translational positive loop where CPEB1 ensues its replacement by CPEB4
Abstract
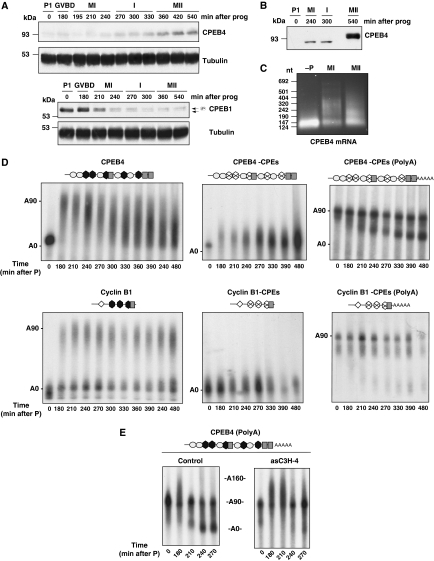
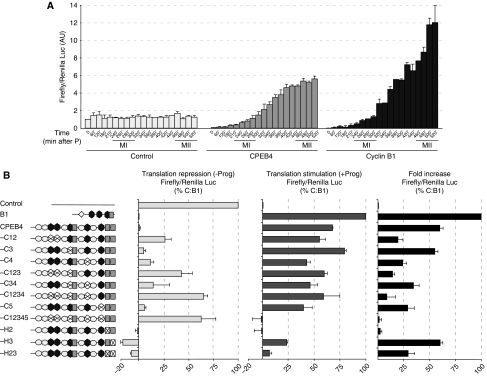
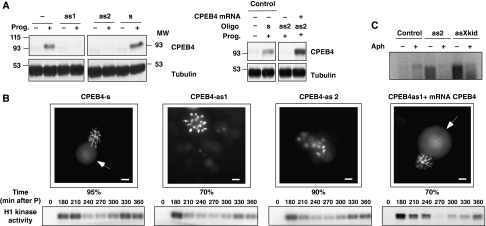
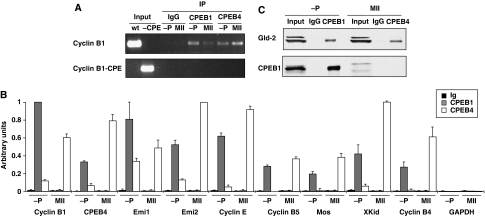
Meiotic progression is driven by the sequential translational activation of maternal messenger RNAs stored in the cytoplasm. This activation is mainly induced by the cytoplasmic elongation of their poly(A) tails, which is mediated by the cytoplasmic polyadenylation element (CPE) present in their 3' untranslated regions. Although polyadenylation in prophase I and metaphase I is mediated by the CPE-binding protein 1 (CPEB1), this protein is degraded during the first meiotic division. Thus, raising the question of how the cytoplasmic polyadenylation required for the second meiotic division is achieved. In this work, we show that CPEB1 generates a positive loop by activating the translation of CPEB4 mRNA, which, in turn, replaces CPEB1 and drives the transition from metaphase I to metaphase II. We further show that CPEB1 and CPEB4 are differentially regulated by phase-specific kinases, generating the need of two sequential CPEB activities to sustain cytoplasmic polyadenylation during all the meiotic phases. Altogether, this work defines a new element in the translational circuit that support an autonomous transition between the two meiotic divisions in the absence of DNA replication.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
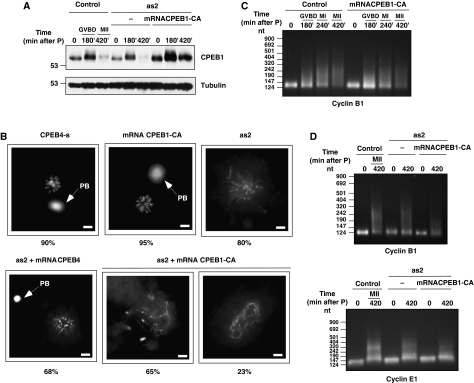
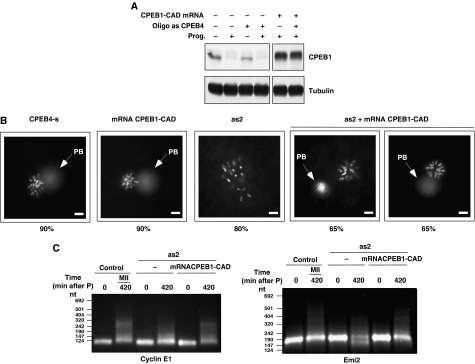
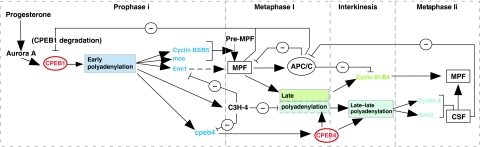
Similar articles
-
A deadenylation negative feedback mechanism governs meiotic metaphase arrest.Nature. 2008 Apr 24;452(7190):1017-21. doi: 10.1038/nature06809. Epub 2008 Apr 2. Nature. 2008. PMID: 18385675
-
Sequential Functions of CPEB1 and CPEB4 Regulate Pathologic Expression of Vascular Endothelial Growth Factor and Angiogenesis in Chronic Liver Disease.Gastroenterology. 2016 Apr;150(4):982-97.e30. doi: 10.1053/j.gastro.2015.11.038. Epub 2015 Nov 26. Gastroenterology. 2016. PMID: 26627607
-
Musashi 1 regulates the timing and extent of meiotic mRNA translational activation by promoting the use of specific CPEs.Nat Struct Mol Biol. 2017 Aug;24(8):672-681. doi: 10.1038/nsmb.3434. Epub 2017 Jul 17. Nat Struct Mol Biol. 2017. PMID: 28714992
-
Translational control by cytoplasmic polyadenylation in Xenopus oocytes.Biochim Biophys Acta. 2008 Apr;1779(4):217-29. doi: 10.1016/j.bbagrm.2008.02.002. Epub 2008 Feb 14. Biochim Biophys Acta. 2008. PMID: 18316045 Free PMC article. Review.
-
Cytoplasmic mRNA polyadenylation and translation assays.Methods Mol Biol. 2006;322:183-98. doi: 10.1007/978-1-59745-000-3_13. Methods Mol Biol. 2006. PMID: 16739724 Review.
Cited by
-
Zar1 represses translation in Xenopus oocytes and binds to the TCS in maternal mRNAs with different characteristics than Zar2.Biochim Biophys Acta. 2013 Oct;1829(10):1034-46. doi: 10.1016/j.bbagrm.2013.06.001. Epub 2013 Jul 1. Biochim Biophys Acta. 2013. PMID: 23827238 Free PMC article.
-
A dynamical model of oocyte maturation unveils precisely orchestrated meiotic decisions.PLoS Comput Biol. 2012 Jan;8(1):e1002329. doi: 10.1371/journal.pcbi.1002329. Epub 2012 Jan 5. PLoS Comput Biol. 2012. PMID: 22238511 Free PMC article.
-
CPEB2, CPEB3 and CPEB4 are coordinately regulated by miRNAs recognizing conserved binding sites in paralog positions of their 3'-UTRs.Nucleic Acids Res. 2010 Nov;38(21):7698-710. doi: 10.1093/nar/gkq635. Epub 2010 Jul 25. Nucleic Acids Res. 2010. PMID: 20660482 Free PMC article.
-
CPEB4 knockout mice exhibit normal hippocampus-related synaptic plasticity and memory.PLoS One. 2013 Dec 30;8(12):e84978. doi: 10.1371/journal.pone.0084978. eCollection 2013. PLoS One. 2013. PMID: 24386439 Free PMC article.
-
Genetic dissection of the planarian reproductive system through characterization of Schmidtea mediterranea CPEB homologs.Dev Biol. 2017 Jun 1;426(1):43-55. doi: 10.1016/j.ydbio.2017.04.008. Epub 2017 Apr 21. Dev Biol. 2017. PMID: 28434803 Free PMC article.
References
-
- Aoki K, Matsumoto K, Tsujimoto M (2003) Xenopus cold-inducible RNA-binding protein 2 interacts with ElrA, the Xenopus homolog of HuR, and inhibits deadenylation of specific mRNAs. J Biol Chem 278: 48491–48497 - PubMed
-
- Belloc E, Mendez R (2008) A deadenylation negative feedback mechanism governs meiotic metaphase arrest. Nature 452: 1017–1021 - PubMed
-
- Belloc E, Pique M, Mendez R (2008) Sequential waves of polyadenylation and deadenylation define a translation circuit that drives meiotic progression. Biochem Soc Trans 36(Part 4): 665–670 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
