

Activation of AMPA receptors in the suprachiasmatic nucleus phase-shifts the mouse circadian clock in vivo and in vitro
- PMID: 20532181
- PMCID: PMC2880614
- DOI: 10.1371/journal.pone.0010951
Activation of AMPA receptors in the suprachiasmatic nucleus phase-shifts the mouse circadian clock in vivo and in vitro
Abstract
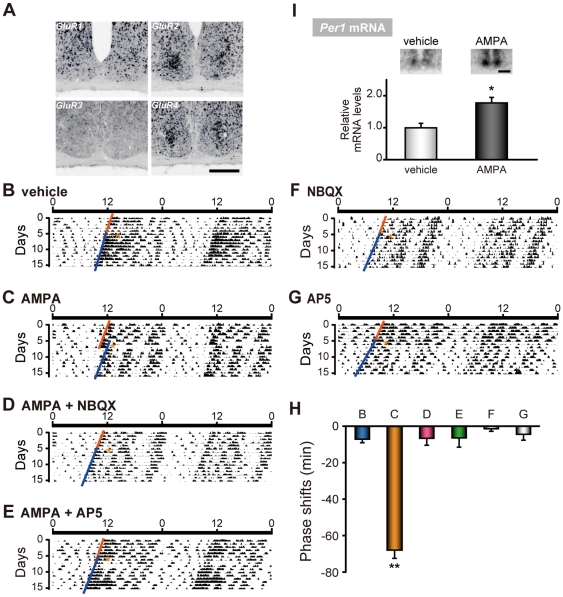
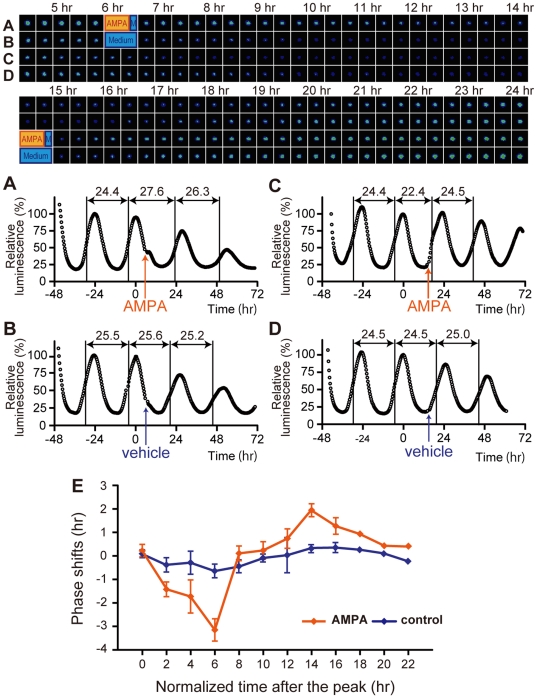
The glutamatergic neurotransmission in the suprachiasmatic nucleus (SCN) plays a central role in the entrainment of the circadian rhythms to environmental light-dark cycles. Although the glutamatergic effect operating via NMDAR (N-methyl D-aspartate receptor) is well elucidated, much less is known about a role of AMPAR (alpha-amino-3-hydroxy-5-methylisoxazole-4-propionic acid receptor) in circadian entrainment. Here we show that, in the mouse SCN, GluR2 and GluR4 AMPAR subtypes are abundantly expressed in the retinorecipient area. In vivo microinjection of AMPA in the SCN during the early subjective night phase-delays the behavioral rhythm. In the organotypic SCN slice culture, AMPA application induces phase-dependent phase-shifts of core-clock gene transcription rhythms. These data demonstrate that activation of AMPAR is capable of phase-shifting the circadian clock both in vivo and in vitro, and are consistent with the hypothesis that activation of AMPA receptors is a critical step in the transmission of photic information to the SCN.
Conflict of interest statement
Figures
Similar articles
-
N-methyl-D-aspartate induces phase shifts in circadian rhythm of neuronal activity of rat SCN in vitro.Am J Physiol. 1994 Aug;267(2 Pt 2):R360-4. doi: 10.1152/ajpregu.1994.267.2.R360. Am J Physiol. 1994. PMID: 7520671
-
Excitatory mechanisms in the suprachiasmatic nucleus: the role of AMPA/KA glutamate receptors.J Neurophysiol. 2002 Aug;88(2):817-28. doi: 10.1152/jn.2002.88.2.817. J Neurophysiol. 2002. PMID: 12163533 Free PMC article.
-
Protein-synthesis inhibitor blocks (R,S)-alpha-amino-3-hydroxy-5-methylisoxazole-4-propionic acid (AMPA)-or substance P-induced phase shift of the circadian rhythm of neuronal activity in the rat suprachiasmatic nucleus in vitro.Neurosci Lett. 1994 Feb 28;168(1-2):159-62. doi: 10.1016/0304-3940(94)90440-5. Neurosci Lett. 1994. PMID: 7518059
-
The role of glutamate in the photic regulation of the suprachiasmatic nucleus.Prog Neurobiol. 1996 Oct;50(2-3):109-32. doi: 10.1016/s0301-0082(96)00032-9. Prog Neurobiol. 1996. PMID: 8971980 Review.
-
Suprachiasmatic nucleus: the brain's circadian clock.Recent Prog Horm Res. 1999;54:33-58; discussion 58-9. Recent Prog Horm Res. 1999. PMID: 10548871 Review.
Cited by
-
Ion Channels Controlling Circadian Rhythms in Suprachiasmatic Nucleus Excitability.Physiol Rev. 2020 Oct 1;100(4):1415-1454. doi: 10.1152/physrev.00027.2019. Epub 2020 Mar 12. Physiol Rev. 2020. PMID: 32163720 Free PMC article. Review.
-
Activation of mGluR1 negatively modulates glutamate-induced phase shifts of the circadian pacemaker in the mouse suprachiasmatic nucleus.Neurobiol Sleep Circadian Rhythms. 2023 Feb 19;14:100089. doi: 10.1016/j.nbscr.2023.100089. eCollection 2023 May. Neurobiol Sleep Circadian Rhythms. 2023. PMID: 36874931 Free PMC article.
-
Orchestration of the circadian clock and its association with Alzheimer's disease: Role of endocannabinoid signaling.Ageing Res Rev. 2022 Jan;73:101533. doi: 10.1016/j.arr.2021.101533. Epub 2021 Nov 26. Ageing Res Rev. 2022. PMID: 34844016 Free PMC article. Review.
-
Circadian Disruptions in the Myshkin Mouse Model of Mania Are Independent of Deficits in Suprachiasmatic Molecular Clock Function.Biol Psychiatry. 2018 Dec 1;84(11):827-837. doi: 10.1016/j.biopsych.2017.04.018. Epub 2017 May 20. Biol Psychiatry. 2018. PMID: 28689605 Free PMC article.
-
Loss of neuropeptide signalling alters temporal expression of mouse suprachiasmatic neuronal state and excitability.Eur J Neurosci. 2024 Dec;60(11):6617-6633. doi: 10.1111/ejn.16590. Epub 2024 Nov 17. Eur J Neurosci. 2024. PMID: 39551976 Free PMC article.
References
-
- Dibner C, Schibler U, Albrecht U. The Mammalian circadian timing system: organization and coordination of central and peripheral clocks. Annu Rev Physiol. 2010;72:517–549. - PubMed
-
- Yamazaki S, Numano R, Abe M, Hida A, Takahashi R, et al. Resetting central and peripheral circadian oscillators in transgenic rats. Science. 2000;288:682–685. - PubMed
-
- Dunlap JC. Molecular bases for circadian clocks. Cell. 1999;96:271–290. - PubMed
-
- Reppert SM, Weaver DR. Molecular analysis of mammalian circadian rhythms. Annu Rev Physiol. 2001;63:647–676. - PubMed
-
- Moore RY, Lenn NJ. A retinohypothalamic projection in the rat. J Comp Neurol. 1972;146:1–14. - PubMed
