

Mice deficient in brain-derived neurotrophic factor have altered development of gastric vagal sensory innervation
- PMID: 20533354
- PMCID: PMC2888090
- DOI: 10.1002/cne.22372
Mice deficient in brain-derived neurotrophic factor have altered development of gastric vagal sensory innervation
Abstract
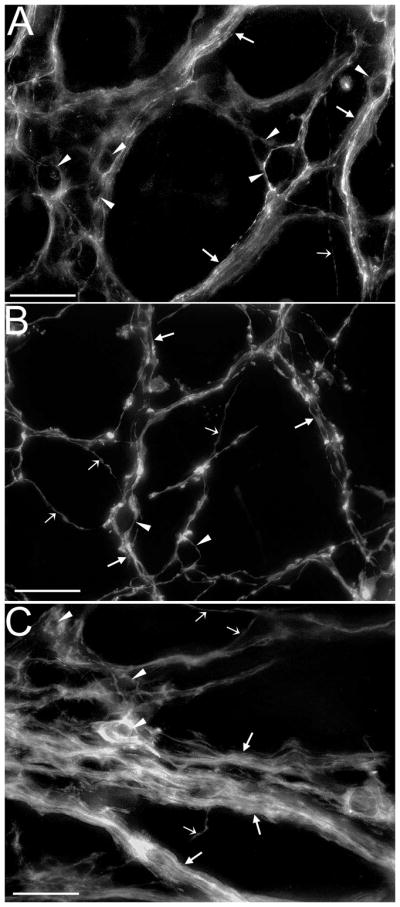
Vagal sensory neurons are dependent on neurotrophins for survival during development. Here, the contribution of brain-derived neurotrophic factor (BDNF) to survival and other aspects of gastric vagal afferent development was investigated. Post-mortem anterograde tracing with 1,1'-dioctadecyl-3,3,3',3'-tetramethylindocarbo-cyanine perchlorate (DiI) was used to label selectively vagal projections to the stomach on postnatal days (P) 0, 3, 4, and 6 in wild types and heterozygous or homozygous BDNF mutants. Sampling sites distributed throughout the ventral stomach wall were scanned with a confocal microscope, and vagal axon bundles, single axons, putative mechanoreceptor precursors (intraganglionic laminar endings, IGLEs; intramuscular arrays, IMAs), and efferent terminals were quantified. Also, myenteric neurons, which are innervated by IGLEs, were stained with cuprolinic blue and counted. Quantitative comparisons across wild-type stomach compartments demonstrated that the adult distribution of IMAs was not present at P0 but began to form by P3-6. Among all the quantified elements, at P0, only IGLE density was significantly different in homozygous mutants compared with wild types, exhibiting a 50% reduction. Also, antrum innervation appeared disorganized, and some putative IMA precursors had truncated telodendria. At P3-6, the effect on IGLEs had recovered, the disorganization of antrum innervation had partially recovered, and some IMA telodendria were still truncated. The present results suggest that gastric IGLEs are among the vagal sensory neurons dependent on BDNF for survival or axon guidance. Alternatively, BDNF deficiency may delay gastric IGLE development. Also, BDNF may contribute to IMA differentiation and patterning of antral vagal innervation.
(c) 2010 Wiley-Liss, Inc.
Figures









Similar articles
-
Factors regulating vagal sensory development: potential role in obesities of developmental origin.Physiol Behav. 2008 Apr 22;94(1):90-104. doi: 10.1016/j.physbeh.2007.11.024. Epub 2007 Nov 22. Physiol Behav. 2008. PMID: 18234244 Free PMC article.
-
Vagal afferent innervation of smooth muscle in the stomach and duodenum of the mouse: morphology and topography.J Comp Neurol. 2000 Dec 18;428(3):558-76. doi: 10.1002/1096-9861(20001218)428:3<558::aid-cne11>3.0.co;2-m. J Comp Neurol. 2000. PMID: 11074451
-
Vagal intraganglionic laminar endings and intramuscular arrays mature at different rates in pre-weanling rat stomach.Auton Neurosci. 2002 Nov 29;102(1-2):13-9. doi: 10.1016/s1566-0702(02)00172-8. Auton Neurosci. 2002. PMID: 12492131
-
A genetic approach for investigating vagal sensory roles in regulation of gastrointestinal function and food intake.Auton Neurosci. 2006 Jun 30;126-127:9-29. doi: 10.1016/j.autneu.2006.03.005. Epub 2006 May 4. Auton Neurosci. 2006. PMID: 16677865 Review.
-
Tension and stretch receptors in gastrointestinal smooth muscle: re-evaluating vagal mechanoreceptor electrophysiology.Brain Res Brain Res Rev. 2000 Nov;34(1-2):1-26. doi: 10.1016/s0165-0173(00)00036-9. Brain Res Brain Res Rev. 2000. PMID: 11086184 Review.
Cited by
-
The gastrointestinal tract microbiome and potential link to Alzheimer's disease.Front Neurol. 2014 Apr 4;5:43. doi: 10.3389/fneur.2014.00043. eCollection 2014. Front Neurol. 2014. PMID: 24772103 Free PMC article. Review. No abstract available.
-
Slit/Robo-mediated chemorepulsion of vagal sensory axons in the fetal gut.Dev Dyn. 2013 Jan;242(1):9-15. doi: 10.1002/dvdy.23898. Epub 2012 Dec 4. Dev Dyn. 2013. PMID: 23161783 Free PMC article.
-
Mechanism of hyperphagia contributing to obesity in brain-derived neurotrophic factor knockout mice.Neuroscience. 2013 Jan 15;229:176-99. doi: 10.1016/j.neuroscience.2012.09.078. Epub 2012 Oct 13. Neuroscience. 2013. PMID: 23069761 Free PMC article.
-
You Are What You (First) Eat.Front Hum Neurosci. 2018 Aug 13;12:323. doi: 10.3389/fnhum.2018.00323. eCollection 2018. Front Hum Neurosci. 2018. PMID: 30150928 Free PMC article. Review.
-
Early postnatal overnutrition: potential roles of gastrointestinal vagal afferents and brain-derived neurotrophic factor.Physiol Behav. 2012 Jun 6;106(3):400-12. doi: 10.1016/j.physbeh.2012.04.002. Physiol Behav. 2012. PMID: 22712064 Free PMC article. Review.
References
-
- Barbacid M. The Trk family of neurotrophin receptors. J Neurobiol. 1994;25(11):1386–1403. - PubMed
-
- Berthoud HR, Kressel M, Raybould HE, Neuhuber WL. Vagal sensors in the rat duodenal mucosa: distribution and structure as revealed by in vivo DiI-tracing. Anat Embryol (Berl) 1995;191(3):203–212. - PubMed
-
- Berthoud HR, Patterson LM, Neumann F, Neuhuber WL. Distribution and structure of vagal afferent intraganglionic laminar endings (IGLEs) in the rat gastrointestinal tract. Anat Embryol. 1997a;195(2):183–191. - PubMed
-
- Berthoud HR, Patterson LM, Willing AE, Mueller K, Neuhuber WL. Capsaicin-resistant vagal afferent fibers in the rat gastrointestinal tract: anatomical identification and functional integrity. Brain Res. 1997b;746(1–2):195–206. - PubMed
-
- Berthoud HR, Powley TL. Vagal afferent innervation of the rat fundic stomach: morphological characterization of the gastric tension receptor. J Comp Neurol. 1992;319(2):261–276. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
