

doi: 10.1002/mabi.201000069.
Analysis of PKR activation using analytical ultracentrifugation
Affiliations
- PMID: 20533534
- PMCID: PMC2926283
- DOI: 10.1002/mabi.201000069
Item in Clipboard
Analysis of PKR activation using analytical ultracentrifugation
Macromol Biosci.
.
Abstract
Protein kinase R (PKR) is a central component of the interferon antiviral defense pathway. Upon binding to dsRNA, PKR undergoes autophosphorylation reactions that activate the kinase, resulting in the inhibition of protein synthesis in virally-infected cells. We have used analytical ultracentrifugation and related biophysical methods to quantitatively characterize the stoichiometries, affinities, and free energy couplings that govern the assembly of the macromolecular complexes in the PKR activation pathway. These studies demonstrate that PKR dimerization play a key role in enzymatic activation and support a model where the role of dsRNA is to bring two or more PKR monomers in close proximity to enhance dimerization.
Figures

PKR domain organization and structure. The N-terminal regulatory domain is comprised of two dsRNA binding motifs, dsRBM1 and dsRBM2 connected by an unstructured linker. Each of these motifs adopts the canonical αβββα fold in the NMR structure of dsRNA binding domain (PDB: 1QU6). In the crystal structure of a complex of the PKR kinase domain and eIF2α (PDB: 2A1A), the kinase domain has the typical bilobal structure observed in other eukaryotic protein kinases and dimerizes via the N-terminal lobe (note, the substrate eIF2α was omitted from the figure for clarity)

Characterization of PKR by analytical ultracentrifugation. A) Sedimentation velocity analysis.[32] Continuous sedimentation coefficient distribution analysis of PKR. Conditions: sample concentration, 0.7 mg/ml; rotor speed, 50,000 RPM; temperature, 20°C; interference optics. B) Sedimentation equilibrium analysis. [32] Conditions: sample concentrations, 0.3, 0.6, 1.0, 1.5 and 2.0 mg/ml; rotor speeds, 18,000 ( ) and 24,000 (□) RPM; temperature, 20°C; interference optics. The dataset was globally fit to monomer-dimer model yielding Kd= 446 μM. The solid lines show the best-fit model and the inset shows the residuals. C) Sedimentation velocity analysis of PKR dsRNA binding domain constructs.[35] Continuous sedimentation coefficient distribution analysis of dsRBD (
) and 24,000 (□) RPM; temperature, 20°C; interference optics. The dataset was globally fit to monomer-dimer model yielding Kd= 446 μM. The solid lines show the best-fit model and the inset shows the residuals. C) Sedimentation velocity analysis of PKR dsRNA binding domain constructs.[35] Continuous sedimentation coefficient distribution analysis of dsRBD ( ) and dsRBM 1 (
) and dsRBM 1 ( ). Conditions: sample concentration, 0.5 mg/ml (dsRBD) and 0.6 mg/ml (dsRBM 1); Rotor speed, 50,000 RPM; temperature, 20°C; interference optics.
). Conditions: sample concentration, 0.5 mg/ml (dsRBD) and 0.6 mg/ml (dsRBM 1); Rotor speed, 50,000 RPM; temperature, 20°C; interference optics.
) and 24,000 (□) RPM; temperature, 20°C; interference optics. The dataset was globally fit to monomer-dimer model yielding Kd= 446 μM. The solid lines show the best-fit model and the inset shows the residuals. C) Sedimentation velocity analysis of PKR dsRNA binding domain constructs.[35] Continuous sedimentation coefficient distribution analysis of dsRBD () and dsRBM 1 (). Conditions: sample concentration, 0.5 mg/ml (dsRBD) and 0.6 mg/ml (dsRBM 1); Rotor speed, 50,000 RPM; temperature, 20°C; interference optics.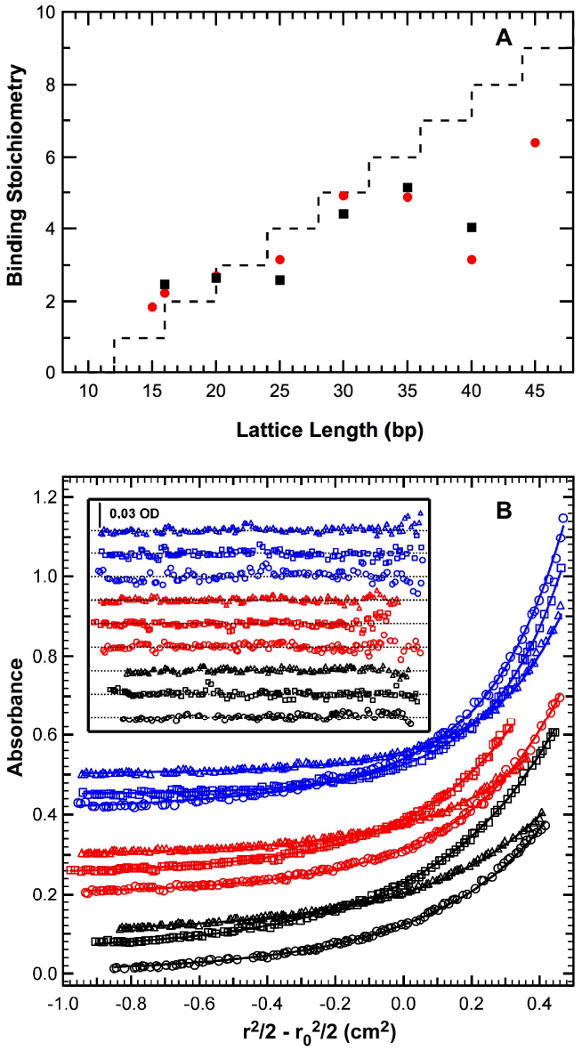
Analysis of the PKR dsRNA binding domain binding to dsRNA. A) Dependence of dsRBD binding stoichiometry on dsRNA length.[35] Stoichiometries were measured by sedimentation equilibrium (black symbols) and CD spectroscopy (red symbols). The dashed line represents the prediction from the overlapping ligand model with N=12 and △=4. B) Multiwavelength sedimentation equilibrium analysis of dsRBD binding to a 20 mer dsRNA.[21] Conditions: rotor speed, 23,000 RPM; temperature, 20°C; RNA concentration, 0.5 μM and protein concentrations of 0.5 μM (black), 1 μM (red) and 2 μM (blue) in 75 mM NaCl, 20 mM HEPES, 5 mM MgCl2, 0.1 mM EDTA, pH 7.5. Detection wavelengths are: 230 nm (), 260 nm (□) and 280 nm (△). Solid lines are a global fit of the data to an unconstrained model of three ligands binding to the 20 mer RNA yielding Kd1= 11 nM, Kd2 = 210 nM and Kd3= 780 nM and an RMSD = 0.00437 OD. Inset: residuals. Traces have been vertically offset for clarity.
), 260 nm (□) and 280 nm (△). Solid lines are a global fit of the data to an unconstrained model of three ligands binding to the 20 mer RNA yielding Kd1= 11 nM, Kd2 = 210 nM and Kd3= 780 nM and an RMSD = 0.00437 OD. Inset: residuals. Traces have been vertically offset for clarity.
Sedimentation velocity analysis of PKR binding to a 20 bp dsRNA.[45] A) Normalized g(s*) distributions of 1 μM dsRNA (black), dsRNA + 0.5 eq. PKR (blue), dsRNA + 1 eq. PKR (red), dsRNA + 2 eq. PKR (green). The distributions are normalized by area. B) Global analysis of sedimentation velocity difference curves. The data were subtracted in pairs and four data sets at the indicated ratios of PKR: dsRNA were fit to 1:1 binding stoichiometry model. The top panels show the data (points) and fit (solid lines) and the bottom panels show the residuals (points). The best fit parameters are shown in Table 1. For clarity, only every 4th difference curve is shown. Conditions: rotor speed, 50,000 RPM; temperature, 20°C; detection: absorption optics at 260 nm; scan interval, 6 minutes.
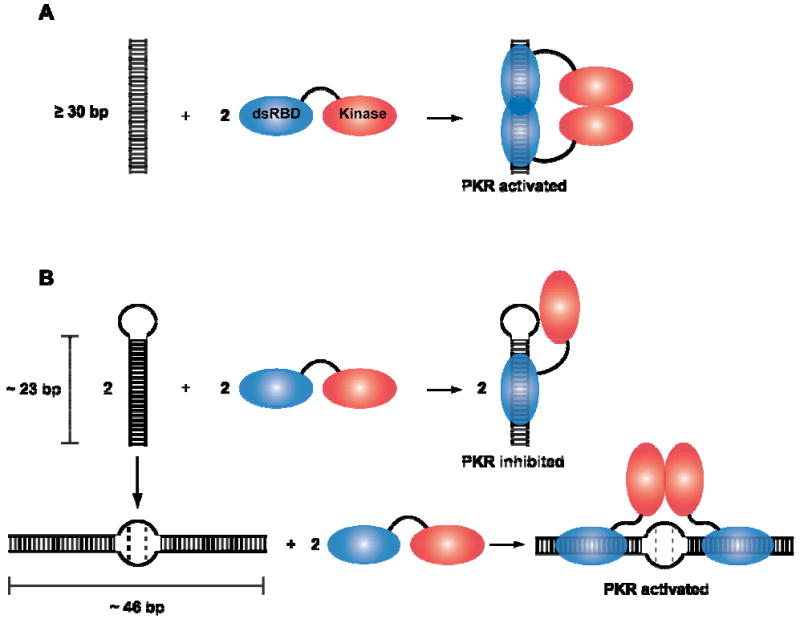
PKR activation models. A) Dimerization model for PKR activation by dsRNA. Binding to dsRNA induces PKR dimerization via the kinase domain, resulting in enzymatic activation. B) Model for PKR activation by TAR RNA dimer. The monomer RNA hairpin (∼23 bp) binds a PKR monomer and fails to activate. In contrast, two PKR monomers can bind to the TAR dimer RNA (∼46 bp) which leads to PKR dimerization and activation.

Simulation of PKR activation by dsRNA. Fraction of PKR present in the RNA-PKR2 species was simulated using a model where two PKR monomers sequentially bind a dsRNA. The binding equilibria were numerically simulated using IGOR Pro (Wavemetrics Inc.) for [PKR] = 200 nM. A ratio of Kd2/Kd1 = 10 was assumed with the following values of Kd1: 1 nM (purple), 3 nM (tan), 10 nM (black), 30 nM (green), 100 nM (blue), 300 nM (red), 1 μM (aqua).

Sedimentation velocity analysis of PKR binding to HIV TAR RNA.[50] A) Secondary structures for TAR variants. Wild-type TAR monomer (wtTAR, left), with changes (red) used to make the single mutants A34U and U37A, and the double mutant A34U:U37A. Wild-type TAR dimer secondary structure (middle) was determined by enzymatic probing experiments.[50] Self-complementary TAR (scTAR, right). This variant deletes the three bulges present in TAR, while retaining the two GU wobbles at positions 6 and 9; in addition, it is mutated on the 3′-side of its loop to add perfect self-complementarity. B) Sedimentation velocity analysis of PKR binding to wtTAR. Top: plot of normalized g(s*) distributions for 1 μM wtTAR monomer (red), 1 μM wtTAR monomer plus 0.5 μM PKR (green), 1 μM PKR (blue) or 4 μM PKR (black). Conditions: temperature, 20°C; rotor speed, 40,000 RPM; detection: absorption optics at 260 nm. Bottom: plot of normalized g(s*) distributions for 0.25 μM wtTAR dimer (red), 0.25 μM wtTAR dimer plus 0.25 μM PKR (green), 0.5 μM PKR (blue) or 1.5 μM PKR (black). Conditions: temperature, 20°C; rotor speed, 40,000 RPM; detection: absorption optics at 260 nm. C) PKR activation assays carried out using native gel-purified wtTAR monomer and dimer. Autophosphorylation reactions were performed at various concentrations of RNA in the presence of 5 μM PKR and were analyzed by SDS-PAGE.
Similar articles
-
Analysis of high-affinity binding of protein kinase R to double-stranded RNA.Biochemistry. 2012 Nov 6;51(44):8764-70. doi: 10.1021/bi301226h. Epub 2012 Oct 26. Biochemistry. 2012. PMID: 23062027 Free PMC article.
-
Activation of PKR by short stem-loop RNAs containing single-stranded arms.RNA. 2016 Jul;22(7):1065-75. doi: 10.1261/rna.053348.115. Epub 2016 May 20. RNA. 2016. PMID: 27208315 Free PMC article.
-
Mechanism of PKR Activation by dsRNA.J Mol Biol. 2008 Aug 29;381(2):351-60. doi: 10.1016/j.jmb.2008.05.056. Epub 2008 May 29. J Mol Biol. 2008. PMID: 18599071 Free PMC article.
-
Activation of PKR: an open and shut case?Trends Biochem Sci. 2007 Feb;32(2):57-62. doi: 10.1016/j.tibs.2006.12.003. Epub 2006 Dec 29. Trends Biochem Sci. 2007. PMID: 17196820 Free PMC article. Review.
-
The regulation of the protein kinase PKR by RNA.Biochimie. 1996;78(11-12):909-14. doi: 10.1016/s0300-9084(97)86712-0. Biochimie. 1996. PMID: 9150867 Review.
Cited by
-
Auto-phosphorylation Represses Protein Kinase R Activity.Sci Rep. 2017 Mar 10;7:44340. doi: 10.1038/srep44340. Sci Rep. 2017. PMID: 28281686 Free PMC article.
-
Heparin activates PKR by inducing dimerization.J Mol Biol. 2011 Nov 11;413(5):973-84. doi: 10.1016/j.jmb.2011.09.025. Epub 2011 Sep 28. J Mol Biol. 2011. PMID: 21978664 Free PMC article.
-
Protein-protein interactions: switch from classical methods to proteomics and bioinformatics-based approaches.Cell Mol Life Sci. 2014 Jan;71(2):205-28. doi: 10.1007/s00018-013-1333-1. Epub 2013 Apr 12. Cell Mol Life Sci. 2014. PMID: 23579629 Free PMC article. Review.
-
Regulation of innate immunity through RNA structure and the protein kinase PKR.Curr Opin Struct Biol. 2011 Feb;21(1):119-27. doi: 10.1016/j.sbi.2010.11.003. Epub 2010 Dec 8. Curr Opin Struct Biol. 2011. PMID: 21145228 Free PMC article. Review.
-
Multi-level regulation of cellular recognition of viral dsRNA.Cell Mol Life Sci. 2013 Jun;70(11):1949-63. doi: 10.1007/s00018-012-1149-4. Epub 2012 Sep 9. Cell Mol Life Sci. 2013. PMID: 22960755 Free PMC article. Review.
References
-
- Kaufman RJ. The double stranded RNA-activated protein kinase PKR. In: Sonenberg N, Hershey JWB, Mathews MB, editors. Translational Control of Gene Expression. Cold Spring Harbor Laboratory Press; Cold Spring Harbor: 2000. p. 503.
-
- Toth AM, Zhang P, Das S, George CX, Samuel CE. Prog Nucleic Acid Res Mol Biol. 2006;81:369. - PubMed
-
- Tian B, Bevilacqua PC, Diegelman-Parente A, Mathews MB. Nature Rev Mol Cell Biol. 2004;5:1013. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
