

Diabetogenic T cells recognize insulin bound to IAg7 in an unexpected, weakly binding register
- PMID: 20534455
- PMCID: PMC2890771
- DOI: 10.1073/pnas.1006545107
Diabetogenic T cells recognize insulin bound to IAg7 in an unexpected, weakly binding register
Abstract
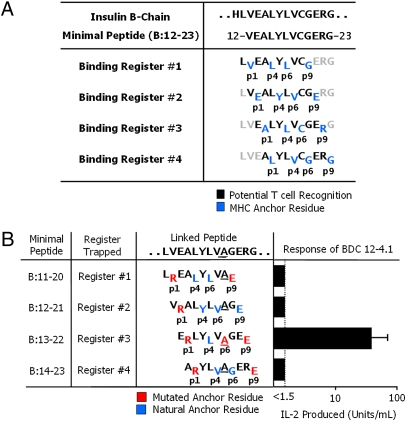
A peptide derived from the insulin B chain contains a major epitope for diabetogenic CD4(+) T cells in the NOD mouse model of type 1 diabetes (T1D). This peptide can fill the binding groove of the NOD MHCII molecule, IA(g7), in a number of ways or "registers." We show here that a diverse set of NOD anti-insulin T cells all recognize this peptide bound in the same register. Surprisingly, this register results in the poorest binding of peptide to IA(g7). The poor binding is due to an incompatibility between the p9 amino acid of the peptide and the unique IA(g7) p9 pocket polymorphisms that are strongly associated with susceptibility to T1D. Our findings suggest that the association of autoimmunity with particular MHCII alleles may be do to poorer, rather than more favorable, binding of the critical self-epitopes, allowing T-cell escape from thymic deletion.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Prochazka M, Leiter EH, Serreze DV, Coleman DL. Three recessive loci required for insulin-dependent diabetes in nonobese diabetic mice. Science. 1987;237:286–289. - PubMed
-
- Todd JA, Bell JI, McDevitt HO. HLA-DQ beta gene contributes to susceptibility and resistance to insulin-dependent diabetes mellitus. Nature. 1987;329:599–604. - PubMed
-
- Reich EP, et al. Self peptides isolated from MHC glycoproteins of non-obese diabetic mice. J Immunol. 1994;152:2279–2288. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- Z01 AI005086/ImNIH/Intramural NIH HHS/United States
- 5 U19-AI05086/AI/NIAID NIH HHS/United States
- R01 DK055969/DK/NIDDK NIH HHS/United States
- AI-18785/AI/NIAID NIH HHS/United States
- R01 AI017134/AI/NIAID NIH HHS/United States
- AI050864/AI/NIAID NIH HHS/United States
- U19 AI050864/AI/NIAID NIH HHS/United States
- R56 AI017134/AI/NIAID NIH HHS/United States
- DK057516/DK/NIDDK NIH HHS/United States
- P01 AI022295/AI/NIAID NIH HHS/United States
- R01 AI018785/AI/NIAID NIH HHS/United States
- P30 DK057516/DK/NIDDK NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- AI-17134/AI/NIAID NIH HHS/United States
- DK55969/DK/NIDDK NIH HHS/United States
- T32 AI007405/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
