

Structure and function of broadly reactive antibody PG16 reveal an H3 subdomain that mediates potent neutralization of HIV-1
- PMID: 20534513
- PMCID: PMC2895122
- DOI: 10.1073/pnas.1004600107
Structure and function of broadly reactive antibody PG16 reveal an H3 subdomain that mediates potent neutralization of HIV-1
Abstract
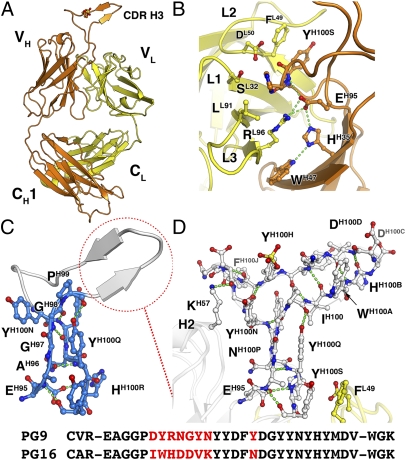
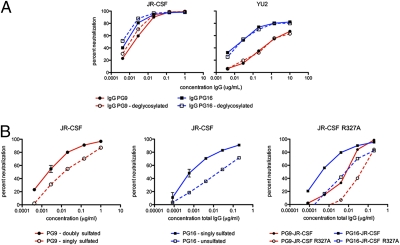
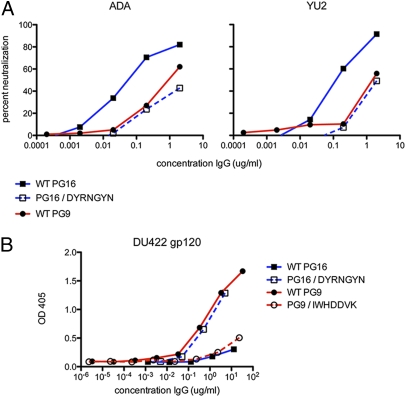
Development of an effective vaccine against HIV-1 will likely require elicitation of broad and potent neutralizing antibodies against the trimeric surface envelope glycoprotein (Env). Monoclonal antibodies (mAbs) PG9 and PG16 neutralize approximately 80% of HIV-1 isolates across all clades with extraordinary potency and target novel epitopes preferentially expressed on Env trimers. As these neutralization properties are ideal for a vaccine-elicited antibody response to HIV-1, their structural basis was investigated. The crystal structure of the antigen-binding fragment (Fab) of PG16 at 2.5 A resolution revealed its unusually long, 28-residue, complementarity determining region (CDR) H3 forms a unique, stable subdomain that towers above the antibody surface. A 7-residue "specificity loop" on the "hammerhead" subdomain was identified that, when transplanted from PG16 to PG9 and vice versa, accounted for differences in the fine specificity and neutralization of these two mAbs. The PG16 electron density maps also revealed that a CDR H3 tyrosine was sulfated, which was confirmed for both PG9 (doubly) and PG16 (singly) by mass spectral analysis. We further showed that tyrosine sulfation plays a role in binding and neutralization. An N-linked glycan modification is observed in the variable light chain, but not required for antigen recognition. Further, the crystal structure of the PG9 light chain at 3.0 A facilitated homology modeling to support the presence of these unusual features in PG9. Thus, PG9 and PG16 use unique structural features to mediate potent neutralization of HIV-1 that may be of utility in antibody engineering and for high-affinity recognition of a variety of therapeutic targets.
Conflict of interest statement
Conflict of interest statement: L.M.W, S.K.P, P.P, and D.R.B. are inventors on a patent describing the human broadly neutralizing antibodies PG9 and PG16 (United States provisional patent application numbers USSN 61/161,010; USSN 61/165,829; and USSN 61/224,739).
Figures
References
-
- Mascola JR, et al. The National Institute of Allergy and Infectious Diseases AIDS Vaccine Evaluation Group Immunization with envelope subunit vaccine products elicits neutralizing antibodies against laboratory-adapted but not primary isolates of human immunodeficiency virus type 1. J Infect Dis. 1996;173:340–348. - PubMed
-
- Flynn NM, et al. rgp120 HIV Vaccine Study Group Placebo-controlled phase 3 trial of a recombinant glycoprotein 120 vaccine to prevent HIV-1 infection. J Infect Dis. 2005;191:654–665. - PubMed
-
- Gilbert PB, et al. Correlation between immunologic responses to a recombinant glycoprotein 120 vaccine and incidence of HIV-1 infection in a phase 3 HIV-1 preventive vaccine trial. J Infect Dis. 2005;191:666–677. - PubMed
-
- Pitisuttithum P, et al. Bangkok Vaccine Evaluation Group Randomized, double-blind, placebo-controlled efficacy trial of a bivalent recombinant glycoprotein 120 HIV-1 vaccine among injection drug users in Bangkok, Thailand. J Infect Dis. 2006;194:1661–1671. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
