

The monoaminergic modulation of sensory-mediated aversive responses in Caenorhabditis elegans requires glutamatergic/peptidergic cotransmission
- PMID: 20534837
- PMCID: PMC3005568
- DOI: 10.1523/JNEUROSCI.0497-10.2010
The monoaminergic modulation of sensory-mediated aversive responses in Caenorhabditis elegans requires glutamatergic/peptidergic cotransmission
Abstract
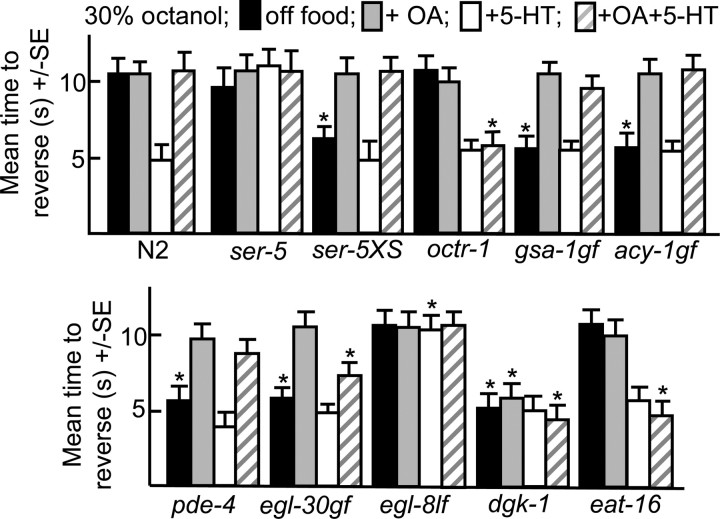
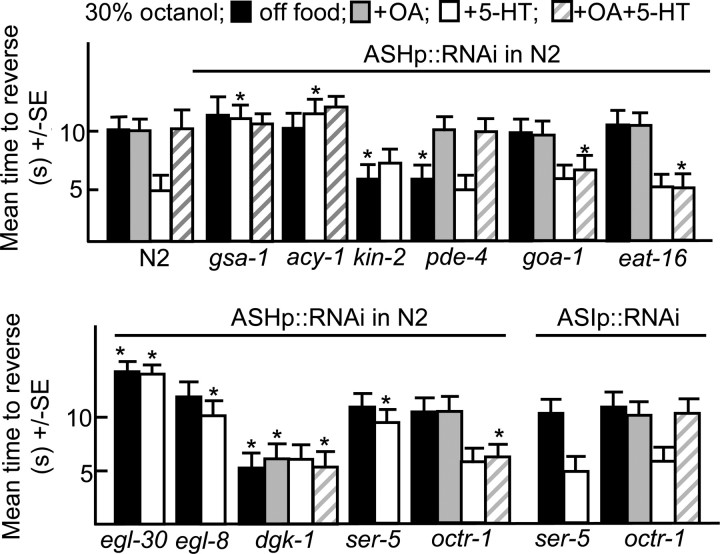
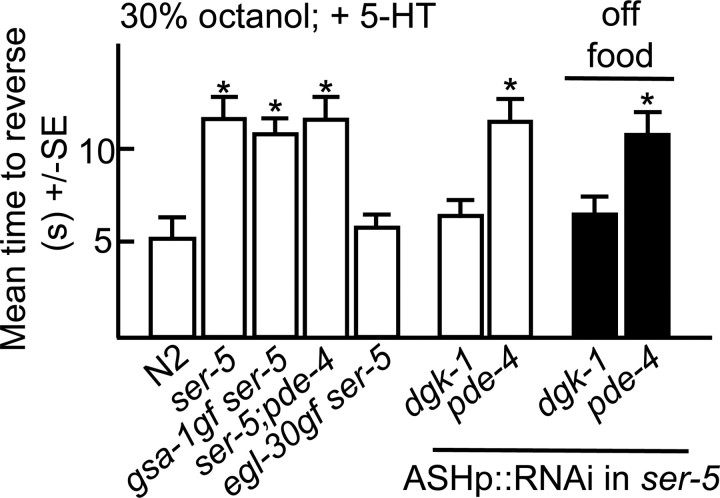
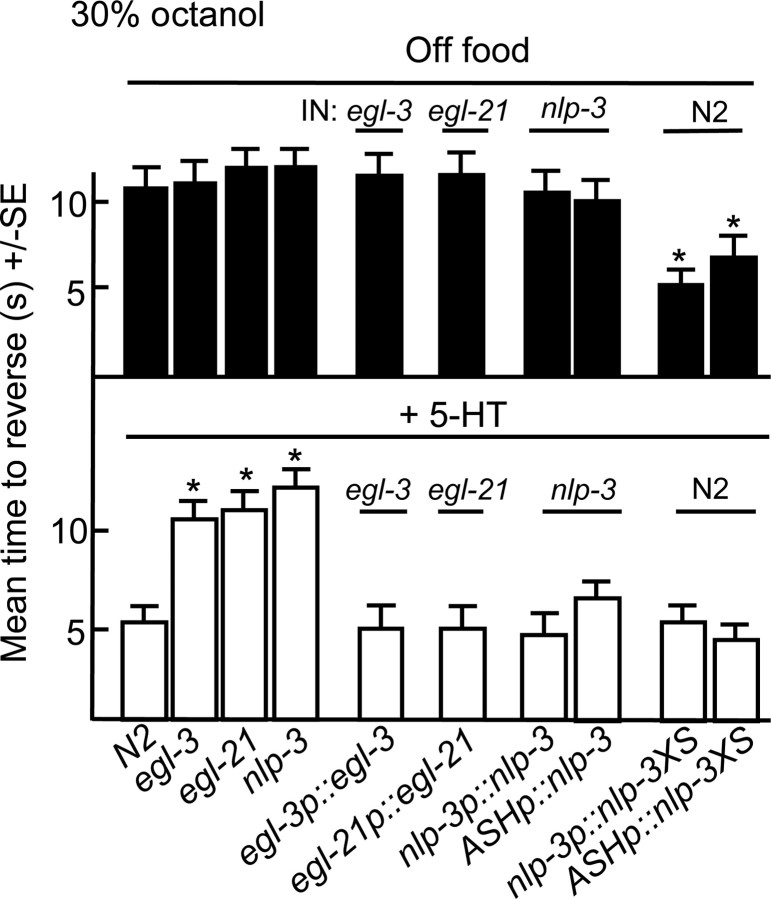
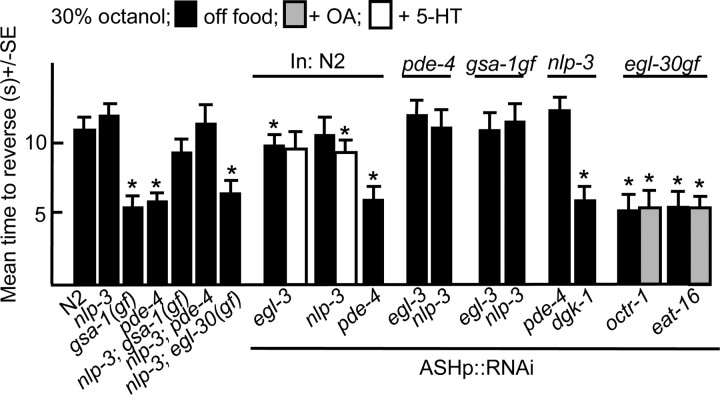
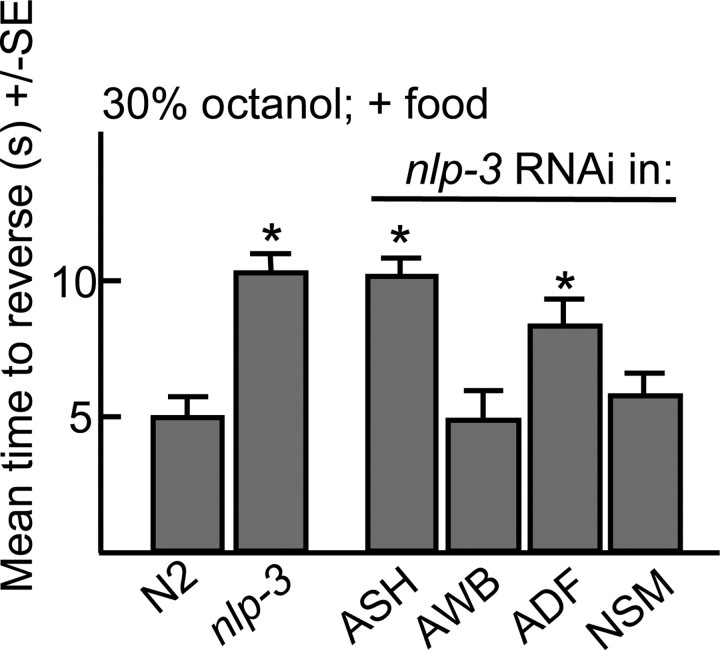
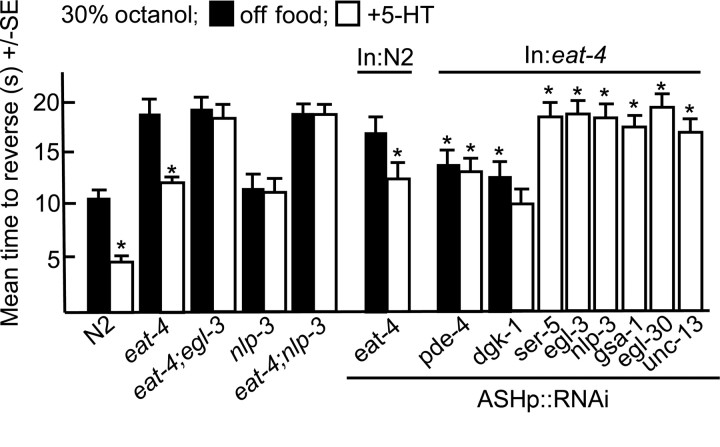
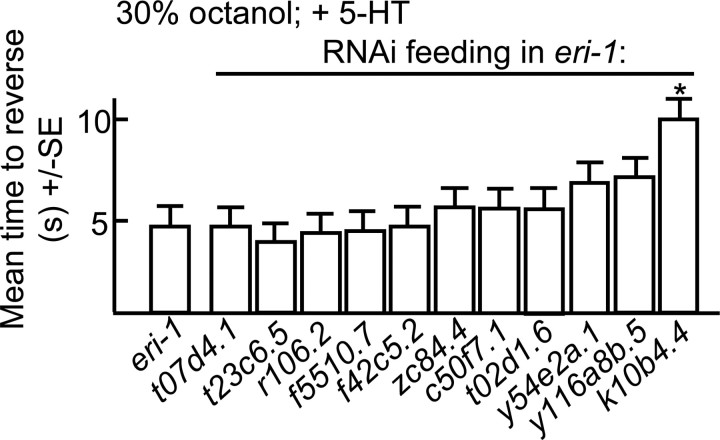
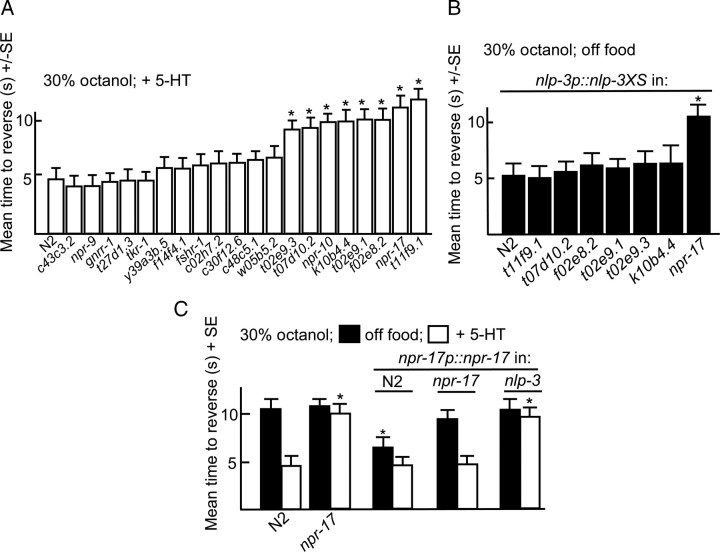
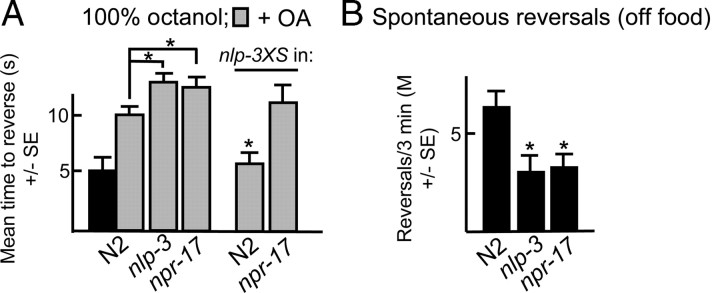
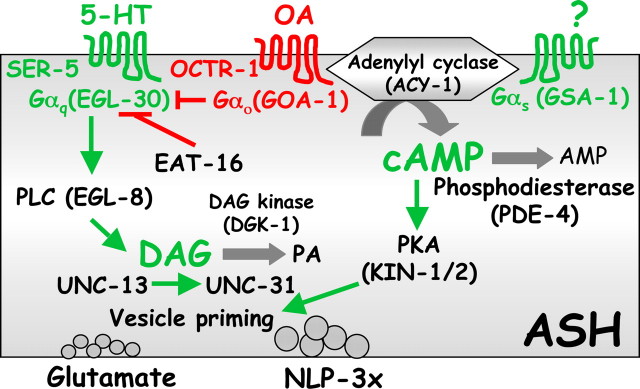
Monoamines and neuropeptides interact to modulate behavioral plasticity in both vertebrates and invertebrates. In Caenorhabditis elegans behavioral state or "mood" is dependent on food availability and is translated by both monoaminergic and peptidergic signaling in the fine-tuning of most behaviors. In the present study, we have examined the interaction of monoamines and peptides on C. elegans aversive behavior mediated by a pair of polymodal, nociceptive, ASH sensory neurons. Food or serotonin sensitize the ASHs and stimulate aversive responses through a pathway requiring the release of nlp-3-encoded neuropeptides from the ASHs. Peptides encoded by nlp-3 appear to stimulate ASH-mediated aversive behavior through the neuropeptide receptor-17 (NPR-17) receptor. nlp-3- and npr-17-null animals exhibit identical phenotypes and animals overexpressing either nlp-3 or npr-17 exhibit elevated aversive responses off food that are absent when nlp-3 or npr-17 are overexpressed in npr-17- or nlp-3-null animals, respectively. ASH-mediated aversive responses are increased by activating either Galpha(q) or Galpha(s) in the ASHs, with Galpha(s) signaling specifically stimulating the release of nlp-3-encoded peptides. In contrast, octopamine appears to inhibit 5-HT stimulation by activating Galpha(o) signaling in the ASHs that, in turn, inhibits both Galpha(s) and Galpha(q) signaling and the release of nlp-3-encoded peptides. These results demonstrate that serotonin and octopamine reversibly modulate the activity of the ASHs, and highlight the utility of the C. elegans model for defining interactions between monoamines and peptides in individual neurons of complex sensory-mediated circuits.
Figures
References
-
- Alkema MJ, Hunter-Ensor M, Ringstad N, Horvitz HR. Tyramine functions independently of octopamine in the Caenorhabditis nervous system. Neuron. 2005;46:247–260. - PubMed
-
- Aspöck G, Ruvkun G, Bürglin TR. The Caenorhabditis elegans ems class homeobox gene ceh-2 is required for M3 pharynx motorneuron function. Development. 2003;130:3369–3378. - PubMed
-
- Bargmann CI, Kaplan JM. Signal transduction in the Caenorhabditis elegans nervous system. Annu Rev Neurosci. 1998;21:279–308. - PubMed
-
- Bargmann CI, Hartwieg E, Horvits HR. Odorant-selective genes and neurons mediate olfaction in C. elegans. Cell. 1993;74:515–527. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous