

Divergent evolution of norovirus GII/4 by genome recombination from May 2006 to February 2009 in Japan
- PMID: 20534859
- PMCID: PMC2916515
- DOI: 10.1128/JVI.02125-09
Divergent evolution of norovirus GII/4 by genome recombination from May 2006 to February 2009 in Japan
Abstract
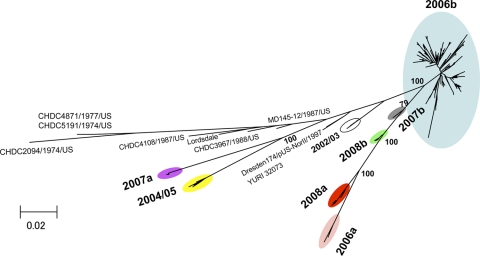
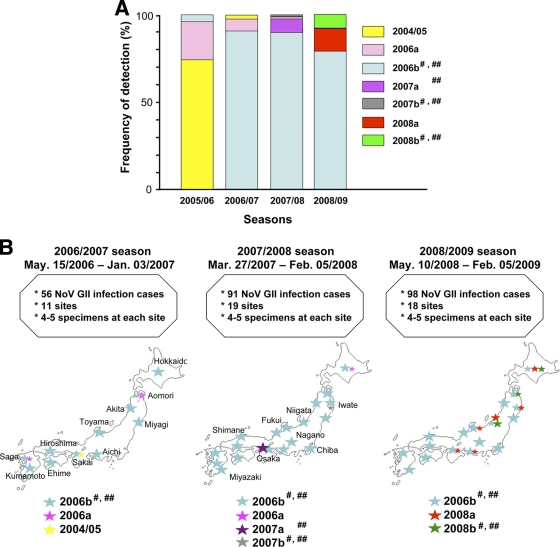
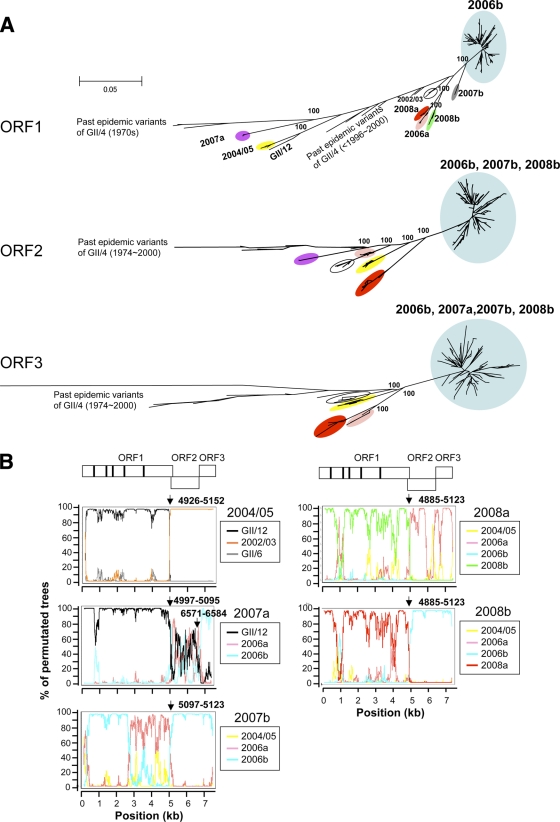
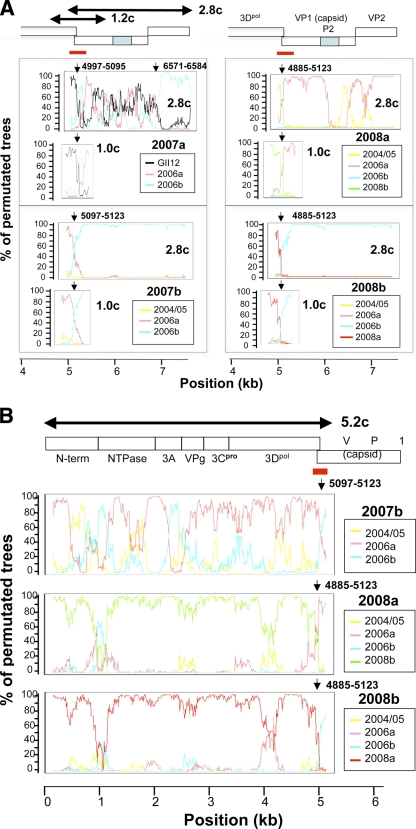
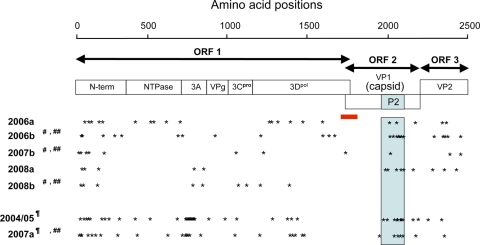
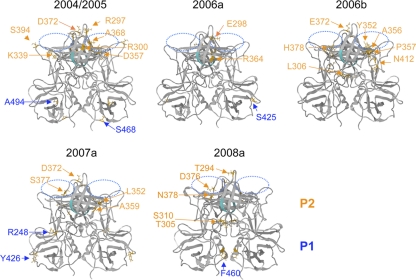
Norovirus GII/4 is a leading cause of acute viral gastroenteritis in humans. We examined here how the GII/4 virus evolves to generate and sustain new epidemics in humans, using 199 near-full-length GII/4 genome sequences and 11 genome segment clones from human stool specimens collected at 19 sites in Japan between May 2006 and February 2009. Phylogenetic studies demonstrated outbreaks of 7 monophyletic GII/4 subtypes, among which a single subtype, termed 2006b, had continually predominated. Phylogenetic-tree, bootscanning-plot, and informative-site analyses revealed that 4 of the 7 GII/4 subtypes were mosaics of recently prevalent GII/4 subtypes and 1 was made up of the GII/4 and GII/12 genotypes. Notably, single putative recombination breakpoints with the highest statistical significance were constantly located around the border of open reading frame 1 (ORF1) and ORF2 (P <or= 0.000001), suggesting outgrowth of specific recombinant viruses in the outbreaks. The GII/4 subtypes had many unique amino acids at the time of their outbreaks, especially in the N-term, 3A-like, and capsid proteins. Unique amino acids in the capsids were preferentially positioned on the outer surface loops of the protruding P2 domain and more abundant in the dominant subtypes. These findings suggest that intersubtype genome recombination at the ORF1/2 boundary region is a common mechanism that realizes independent and concurrent changes on the virion surface and in viral replication proteins for the persistence of norovirus GII/4 in human populations.
Figures
References
-
- Ando, T., J. S. Noel, and R. L. Fankhauser. 2000. Genetic classification of “Norwalk-like viruses.” J. Infect. Dis. 181(Suppl. 2):S336-S348. - PubMed
-
- Baker, D., and A. Sali. 2001. Protein structure prediction and structural genomics. Science 294:93-96. - PubMed
-
- Belliot, G., S. V. Sosnovtsev, T. Mitra, C. Hammer, M. Garfield, and K. Y. Green. 2003. In vitro proteolytic processing of the MD145 norovirus ORF1 nonstructural polyprotein yields stable precursors and products similar to those detected in calicivirus-infected cells. J. Virol. 77:10957-10974. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
