

Loss of enigma homolog protein results in dilated cardiomyopathy
- PMID: 20538684
- PMCID: PMC3684396
- DOI: 10.1161/CIRCRESAHA.110.218735
Loss of enigma homolog protein results in dilated cardiomyopathy
Abstract
Rationale: The Z-line, alternatively termed the Z-band or Z-disc, is a highly ordered structure at the border between 2 sarcomeres. Enigma subfamily proteins (Enigma, Enigma homolog protein, and Cypher) of the PDZ-LIM domain protein family are Z-line proteins. Among the Enigma subfamily, Cypher has been demonstrated to play a pivotal role in the structure and function of striated muscle, whereas the role of Enigma homolog protein (ENH) in muscle remains largely unknown.
Objective: We studied the role of Enigma homolog protein in the heart using global and cardiac-specific ENH knockout mouse models.
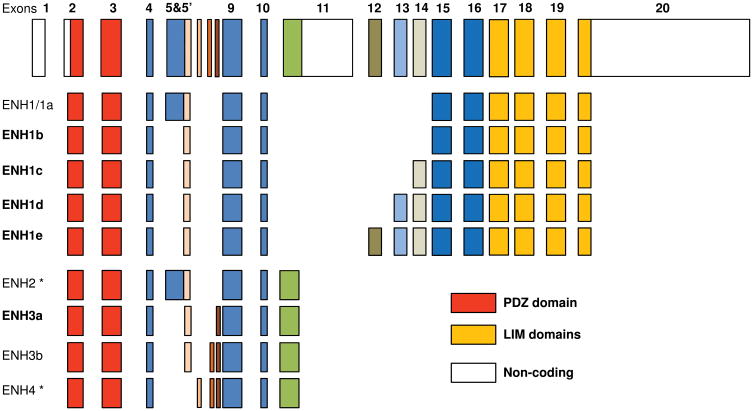
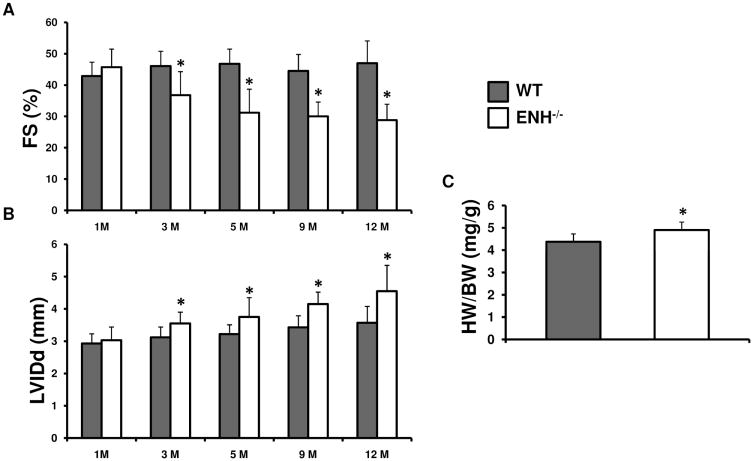
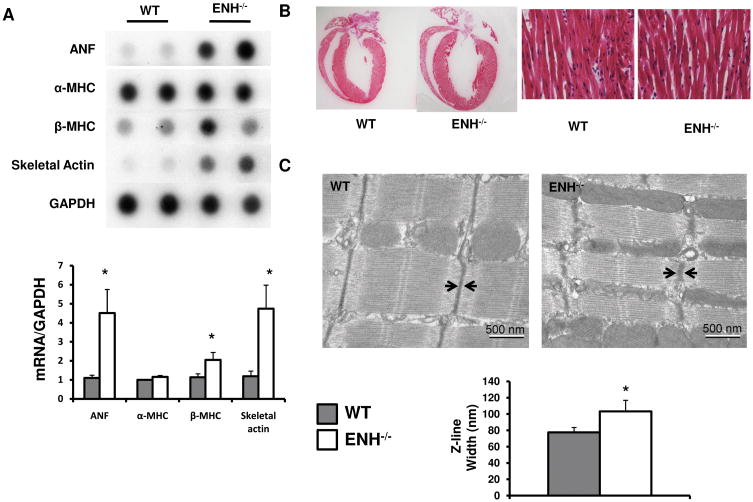
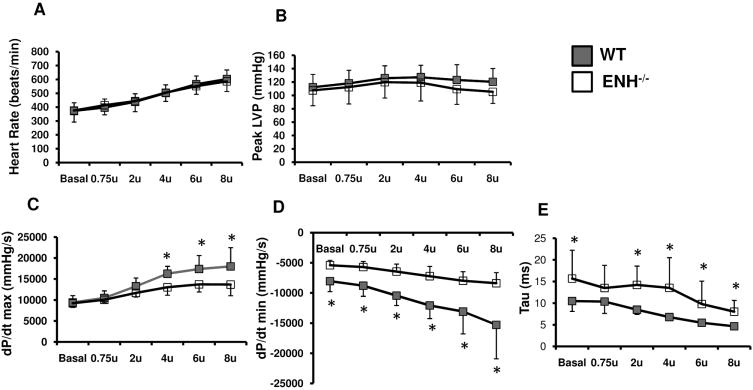
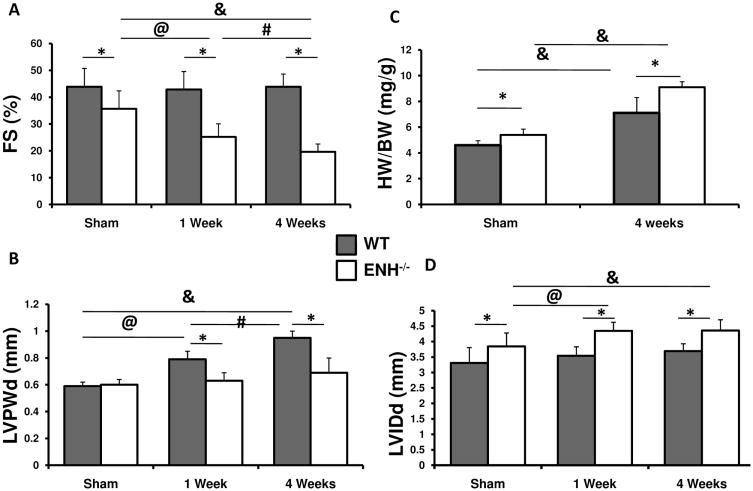
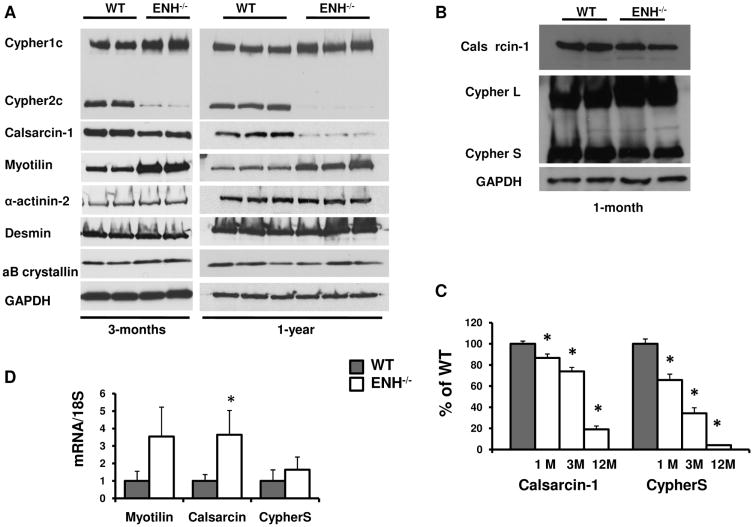
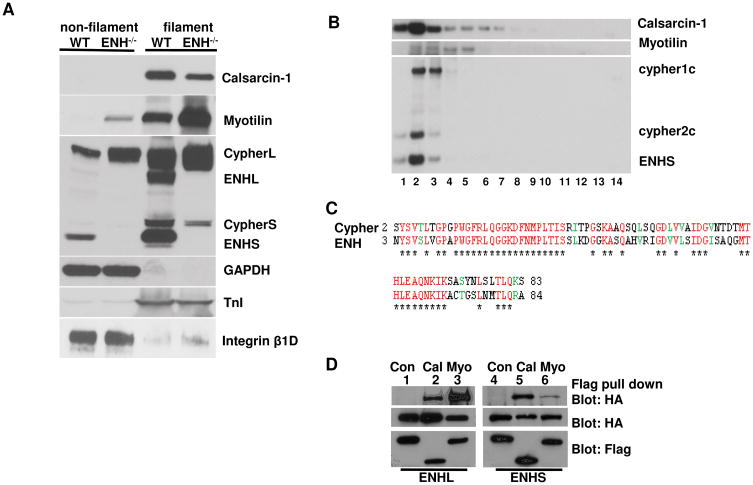
Methods and results: We identified new exons and splice isoforms for ENH in the mouse heart. Impaired cardiac contraction and dilated cardiomyopathy were observed in ENH null mice. Mice with cardiac specific ENH deletion developed a similar dilated cardiomyopathy. Like Cypher, ENH interacted with Calsarcin-1, another Z-line protein. Moreover, biochemical studies showed that ENH, Cypher short isoform and Calsarcin-1 are within the same protein complex at the Z-line. Cypher short isoform and Calsarcin-1 proteins are specifically downregulated in ENH null hearts.
Conclusions: We have identified an ENH-CypherS-Calsarcin protein complex at the Z-line. Ablation of ENH leads to destabilization of this protein complex and dilated cardiomyopathy.
Figures
Comment in
-
Unraveling enigma in the z-disks.Circ Res. 2010 Aug 6;107(3):321-3. doi: 10.1161/CIRCRESAHA.110.225615. Circ Res. 2010. PMID: 20689070 No abstract available.
Similar articles
-
Cypher and Enigma homolog protein are essential for cardiac development and embryonic survival.J Am Heart Assoc. 2015 May 5;4(5):e001950. doi: 10.1161/JAHA.115.001950. J Am Heart Assoc. 2015. PMID: 25944877 Free PMC article.
-
Cardiac-specific ablation of Cypher leads to a severe form of dilated cardiomyopathy with premature death.Hum Mol Genet. 2009 Feb 15;18(4):701-13. doi: 10.1093/hmg/ddn400. Epub 2008 Nov 21. Hum Mol Genet. 2009. PMID: 19028670 Free PMC article.
-
Selective deletion of long but not short Cypher isoforms leads to late-onset dilated cardiomyopathy.Hum Mol Genet. 2011 May 1;20(9):1751-62. doi: 10.1093/hmg/ddr050. Epub 2011 Feb 8. Hum Mol Genet. 2011. PMID: 21303826 Free PMC article.
-
"Z"eroing in on the role of Cypher in striated muscle function, signaling, and human disease.Trends Cardiovasc Med. 2007 Nov;17(8):258-62. doi: 10.1016/j.tcm.2007.09.002. Trends Cardiovasc Med. 2007. PMID: 18021935 Free PMC article. Review.
-
Roles of Nebulin Family Members in the Heart.Circ J. 2015;79(10):2081-7. doi: 10.1253/circj.CJ-15-0854. Epub 2015 Aug 31. Circ J. 2015. PMID: 26321576 Review.
Cited by
-
Extracellular vesicles in cardiomyopathies: A narrative review.Heliyon. 2023 Dec 17;10(1):e23765. doi: 10.1016/j.heliyon.2023.e23765. eCollection 2024 Jan 15. Heliyon. 2023. PMID: 38192847 Free PMC article. Review.
-
The Role of Leucine-Rich Repeat Containing Protein 10 (LRRC10) in Dilated Cardiomyopathy.Front Physiol. 2016 Aug 3;7:337. doi: 10.3389/fphys.2016.00337. eCollection 2016. Front Physiol. 2016. PMID: 27536250 Free PMC article. Review.
-
PDLIM5 inhibits STUB1-mediated degradation of SMAD3 and promotes the migration and invasion of lung cancer cells.J Biol Chem. 2020 Oct 2;295(40):13798-13811. doi: 10.1074/jbc.RA120.014976. Epub 2020 Jul 31. J Biol Chem. 2020. PMID: 32737199 Free PMC article.
-
Leveraging Extracellular Non-coding RNAs to Diagnose and Treat Heart Diseases.J Cardiovasc Transl Res. 2022 Jun;15(3):456-468. doi: 10.1007/s12265-022-10252-x. Epub 2022 Apr 13. J Cardiovasc Transl Res. 2022. PMID: 35419773 Review.
-
LRRC10 is required to maintain cardiac function in response to pressure overload.Am J Physiol Heart Circ Physiol. 2016 Jan 15;310(2):H269-78. doi: 10.1152/ajpheart.00717.2014. Epub 2015 Nov 25. Am J Physiol Heart Circ Physiol. 2016. PMID: 26608339 Free PMC article.
References
-
- Sanger JM, Sanger JW. The dynamic z bands of striated muscle cells. Sci Signal. 2008;1:pe37. - PubMed
-
- Goldstein MA, Schroeter JP, Michael LH. Role of the z band in the mechanical properties of the heart. FASEB J. 1991;5:2167–2174. - PubMed
-
- Towbin JA, Bowles NE. The failing heart. Nature. 2002;415:227–233. - PubMed
-
- Ervasti JM. Costameres: The achilles' heel of herculean muscle. J Biol Chem. 2003;278:13591–13594. - PubMed
-
- Kruger M, Linke WA. Titin-based mechanical signalling in normal and failing myocardium. J Mol Cell Cardiol. 2009;46:490–498. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
