

Adrenergic receptor-mediated disinhibition of mitral cells triggers long-term enhancement of synchronized oscillations in the olfactory bulb
- PMID: 20538781
- PMCID: PMC2934928
- DOI: 10.1152/jn.00328.2010
Adrenergic receptor-mediated disinhibition of mitral cells triggers long-term enhancement of synchronized oscillations in the olfactory bulb
Abstract
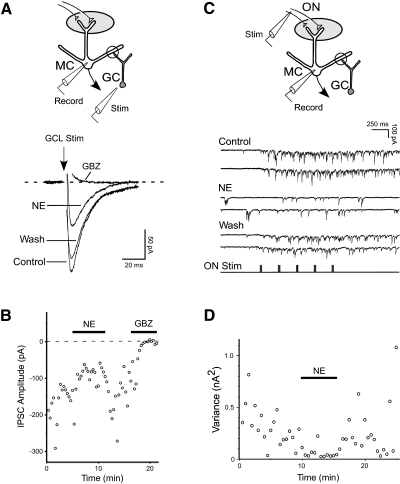
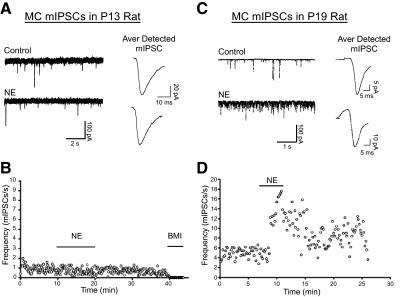
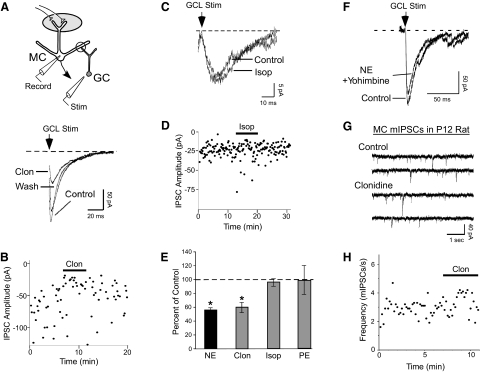
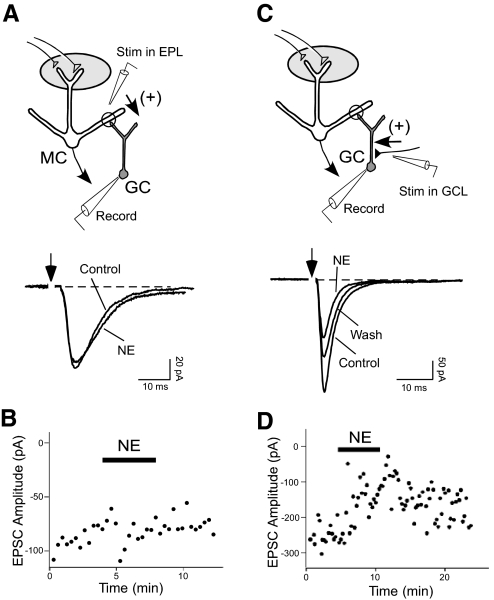
Norepinephrine (NE) is widely implicated in various forms of associative olfactory learning in rodents, including early learning preference in neonates. Here we used patch-clamp recordings in rat olfactory bulb slices to assess cellular actions of NE, examining both acute, short-term effects of NE as well as the relationship between these acute effects and long-term cellular changes that could underlie learning. Our focus for long-term effects was on synchronized gamma frequency (30-70 Hz) oscillations, shown in prior studies to be enhanced for up to an hour after brief exposure of a bulb slice to NE and neuronal stimulation. In terms of acute effects, we found that a dominant action of NE was to reduce inhibitory GABAergic transmission from granule cells (GCs) to output mitral cells (MCs). This disinhibition was also induced by clonidine, an agonist specific for alpha(2) adrenergic receptors (ARs). Acute NE-induced disinhibition of MCs appeared to be linked to long-term enhancement of gamma oscillations, based, first, on the fact that clonidine, but not agonists specific for other AR subtypes, mimicked NE's long-term actions. In addition, the alpha(2) AR-specific antagonist yohimbine blocked the long-term enhancement of the oscillations due to NE. Last, brief exposure of the slice to the GABA(A) receptor antagonist gabazine, to block inhibitory synapses directly, also induced the long-term changes. Acute disinhibition is a plausible permissive effect of NE leading to olfactory learning, because, when combined with exposure to a specific odor, it should lead to neuron-specific increases in intracellular calcium of the type generally associated with long-term synaptic modifications.
Figures
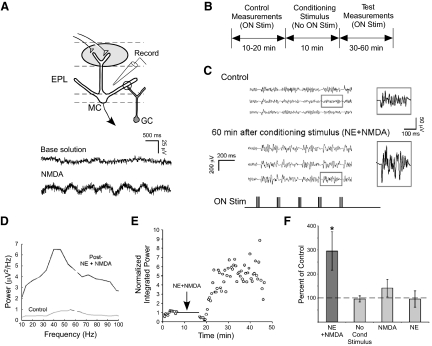
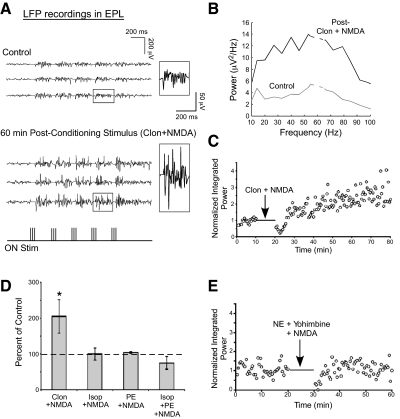

Similar articles
-
α(1A)-Adrenergic regulation of inhibition in the olfactory bulb.J Physiol. 2013 Apr 1;591(7):1631-43. doi: 10.1113/jphysiol.2012.248591. Epub 2012 Dec 24. J Physiol. 2013. PMID: 23266935 Free PMC article.
-
Long-term enhancement of synchronized oscillations by adrenergic receptor activation in the olfactory bulb.J Neurophysiol. 2008 Apr;99(4):2021-5. doi: 10.1152/jn.01324.2007. Epub 2008 Feb 6. J Neurophysiol. 2008. PMID: 18256160
-
Age-dependent adrenergic actions in the main olfactory bulb that could underlie an olfactory-sensitive period.J Neurophysiol. 2012 Oct;108(7):1999-2007. doi: 10.1152/jn.00322.2012. Epub 2012 Jul 18. J Neurophysiol. 2012. PMID: 22815401 Free PMC article.
-
Interplay between local GABAergic interneurons and relay neurons generates gamma oscillations in the rat olfactory bulb.J Neurosci. 2004 May 5;24(18):4382-92. doi: 10.1523/JNEUROSCI.5570-03.2004. J Neurosci. 2004. PMID: 15128852 Free PMC article.
-
Direct excitation of mitral cells via activation of alpha1-noradrenergic receptors in rat olfactory bulb slices.J Neurophysiol. 2001 Nov;86(5):2173-82. doi: 10.1152/jn.2001.86.5.2173. J Neurophysiol. 2001. PMID: 11698509
Cited by
-
Cortical feedback and gating in odor discrimination and generalization.PLoS Comput Biol. 2021 Oct 11;17(10):e1009479. doi: 10.1371/journal.pcbi.1009479. eCollection 2021 Oct. PLoS Comput Biol. 2021. PMID: 34634035 Free PMC article.
-
Locus Coeruleus Activation Patterns Differentially Modulate Odor Discrimination Learning and Odor Valence in Rats.Cereb Cortex Commun. 2021 Apr 5;2(2):tgab026. doi: 10.1093/texcom/tgab026. eCollection 2021. Cereb Cortex Commun. 2021. PMID: 34296171 Free PMC article.
-
Complex neural representation of odour information in the olfactory bulb.Acta Physiol (Oxf). 2020 Jan;228(1):e13333. doi: 10.1111/apha.13333. Epub 2019 Jul 2. Acta Physiol (Oxf). 2020. PMID: 31188539 Free PMC article. Review.
-
Neural correlates of olfactory learning: Critical role of centrifugal neuromodulation.Learn Mem. 2010 Oct 27;17(11):561-70. doi: 10.1101/lm.941510. Print 2010 Nov. Learn Mem. 2010. PMID: 20980444 Free PMC article.
-
α(1A)-Adrenergic regulation of inhibition in the olfactory bulb.J Physiol. 2013 Apr 1;591(7):1631-43. doi: 10.1113/jphysiol.2012.248591. Epub 2012 Dec 24. J Physiol. 2013. PMID: 23266935 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials