

The PB2 subunit of the influenza virus RNA polymerase affects virulence by interacting with the mitochondrial antiviral signaling protein and inhibiting expression of beta interferon
- PMID: 20538852
- PMCID: PMC2919034
- DOI: 10.1128/JVI.00879-10
The PB2 subunit of the influenza virus RNA polymerase affects virulence by interacting with the mitochondrial antiviral signaling protein and inhibiting expression of beta interferon
Abstract
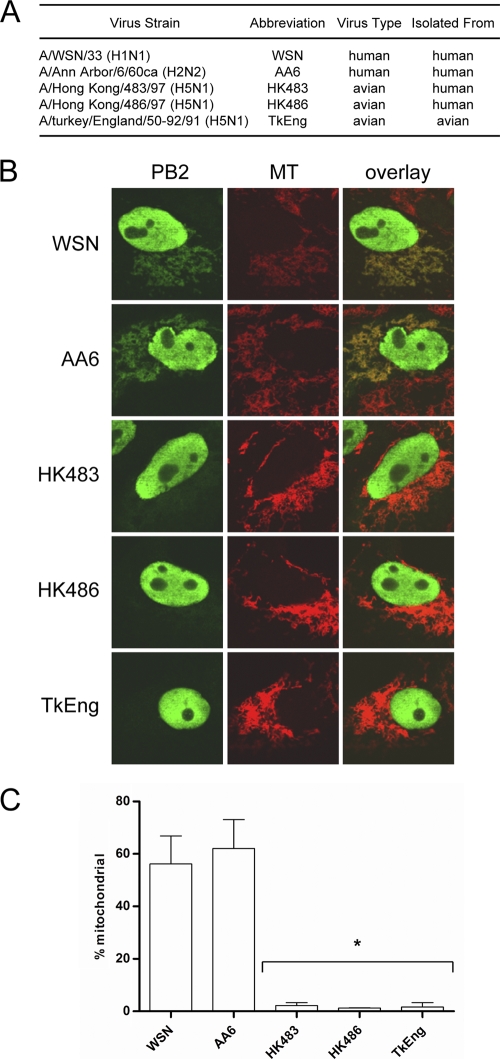
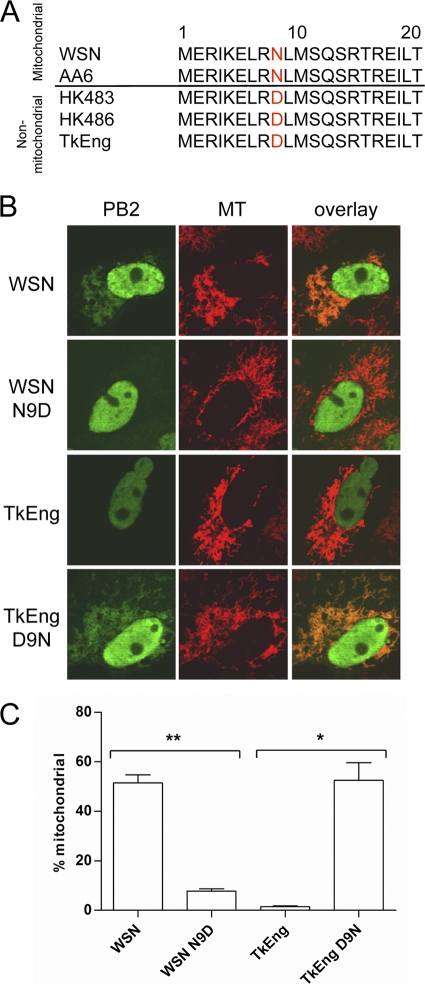
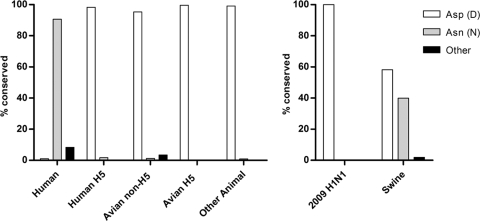
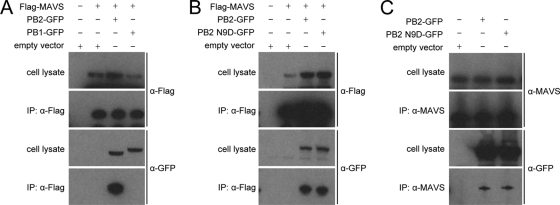
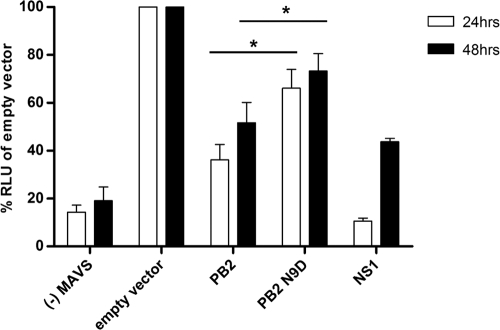
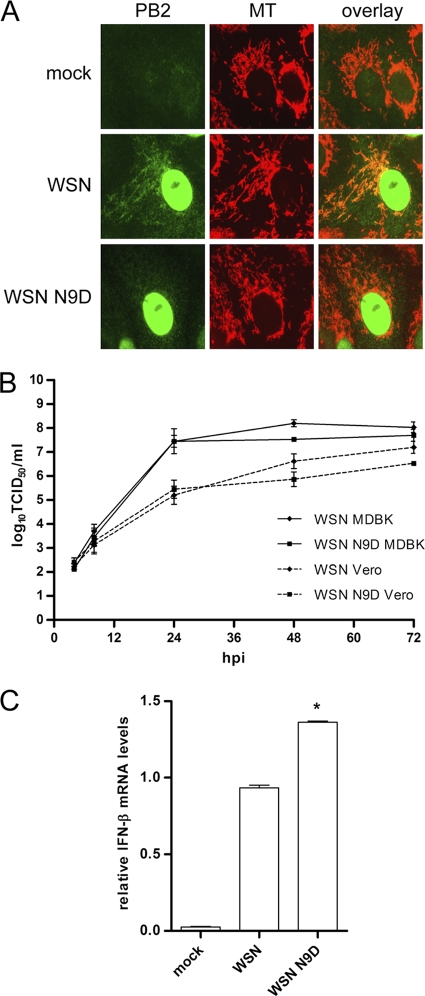
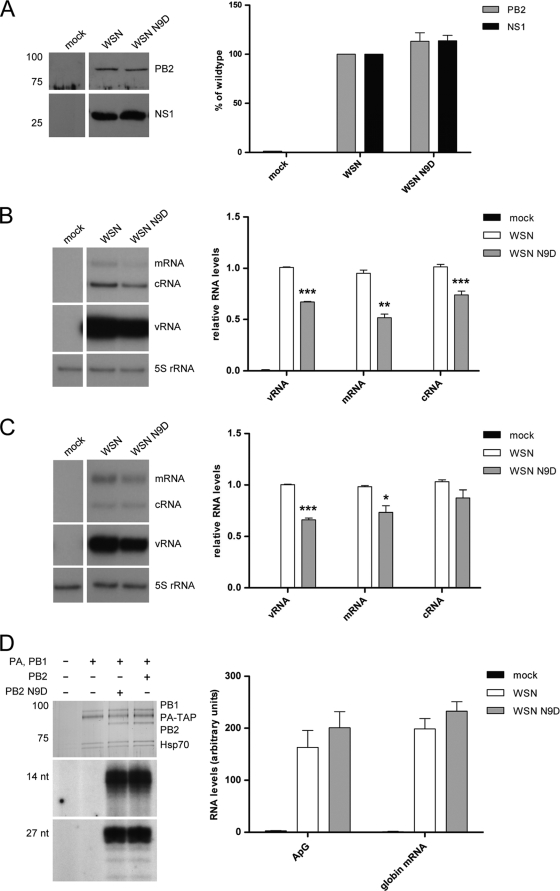
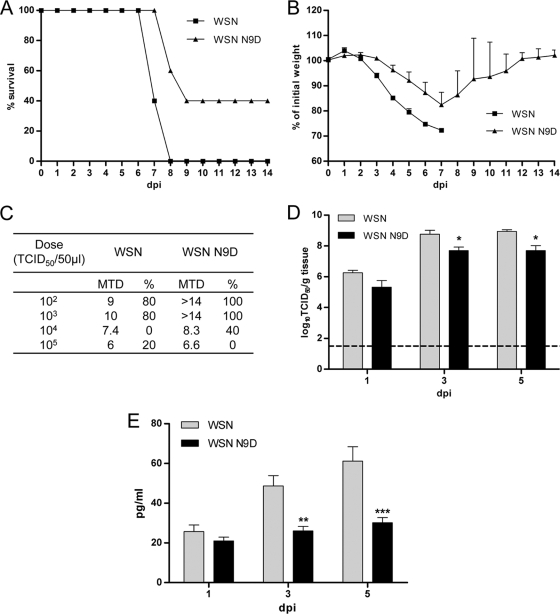
The PB2 subunit of the influenza virus RNA polymerase is a major virulence determinant of influenza viruses. However, the molecular mechanisms involved remain unknown. It was previously shown that the PB2 protein, in addition to its nuclear localization, also accumulates in the mitochondria. Here, we demonstrate that the PB2 protein interacts with the mitochondrial antiviral signaling protein, MAVS (also known as IPS-1, VISA, or Cardif), and inhibits MAVS-mediated beta interferon (IFN-beta) expression. In addition, we show that PB2 proteins of influenza viruses differ in their abilities to associate with the mitochondria. In particular, the PB2 proteins of seasonal human influenza viruses localize to the mitochondria while PB2 proteins of avian influenza viruses are nonmitochondrial. This difference in localization is caused by a single amino acid polymorphism in the PB2 mitochondrial targeting signal. In order to address the functional significance of the mitochondrial localization of the PB2 protein in vivo, we have generated two recombinant human influenza viruses encoding either mitochondrial or nonmitochondrial PB2 proteins. We found that the difference in the mitochondrial localization of the PB2 proteins does not affect the growth of these viruses in cell culture. However, the virus encoding the nonmitochondrial PB2 protein induces higher levels of IFN-beta and, in an animal model, is attenuated compared to the isogenic virus encoding a mitochondrial PB2. Overall this study implicates the PB2 protein in the regulation of host antiviral innate immune pathways and suggests an important role for the mitochondrial association of the PB2 protein in determining virulence.
Figures
References
-
- Almond, J. W. 1977. A single gene determines the host range of influenza virus. Nature 270:617-618. - PubMed
-
- Carr, S. M., E. Carnero, A. Garcia-Sastre, G. G. Brownlee, and E. Fodor. 2006. Characterization of a mitochondrial-targeting signal in the PB2 protein of influenza viruses. Virology 344:492-508. - PubMed
-
- Chen, H., R. A. Bright, K. Subbarao, C. Smith, N. J. Cox, J. M. Katz, and Y. Matsuoka. 2007. Polygenic virulence factors involved in pathogenesis of 1997 Hong Kong H5N1 influenza viruses in mice. Virus Res. 128:159-163. - PubMed
-
- Chen, W., P. A. Calvo, D. Malide, J. Gibbs, U. Schubert, I. Bacik, S. Basta, R. O'Neill, J. Schickli, P. Palese, P. Henklein, J. R. Bennink, and J. W. Yewdell. 2001. A novel influenza A virus mitochondrial protein that induces cell death. Nat. Med. 7:1306-1312. - PubMed
-
- Claas, E. C., A. D. Osterhaus, R. van Beek, J. C. De Jong, G. F. Rimmelzwaan, D. A. Senne, S. Krauss, K. F. Shortridge, and R. G. Webster. 1998. Human influenza A H5N1 virus related to a highly pathogenic avian influenza virus. Lancet 351:472-477. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous

