

Cell cycle progression or translation control is not essential for vesicular stomatitis virus oncolysis of hepatocellular carcinoma
- PMID: 20539760
- PMCID: PMC2881869
- DOI: 10.1371/journal.pone.0010988
Cell cycle progression or translation control is not essential for vesicular stomatitis virus oncolysis of hepatocellular carcinoma
Abstract
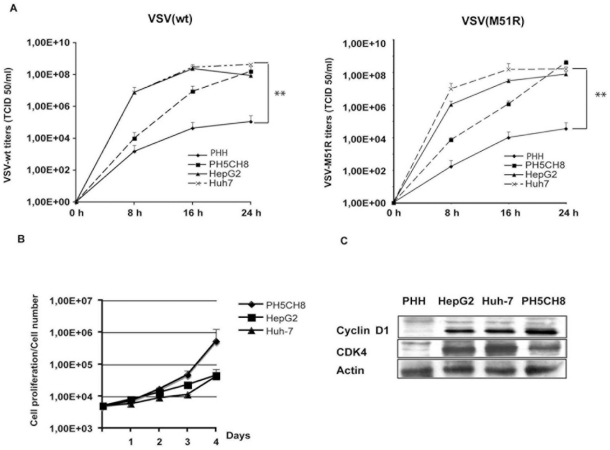
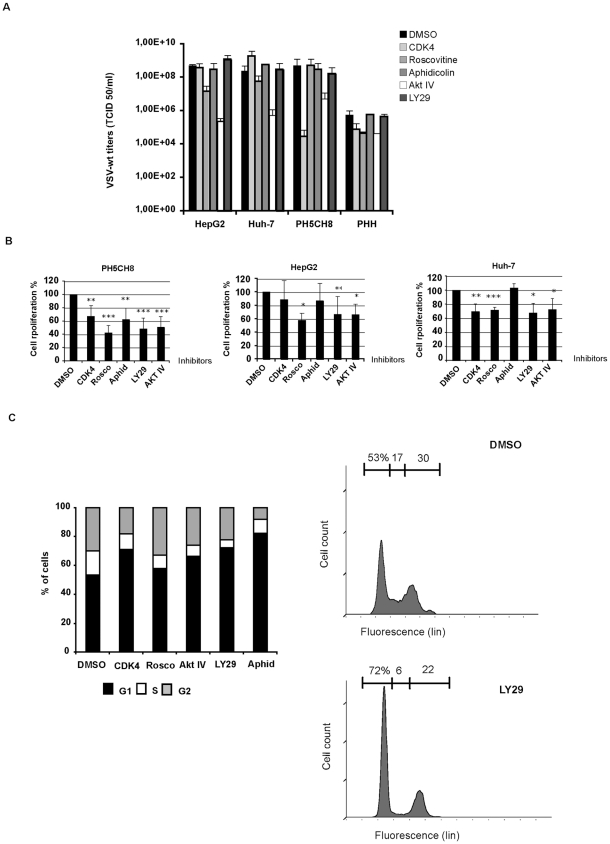
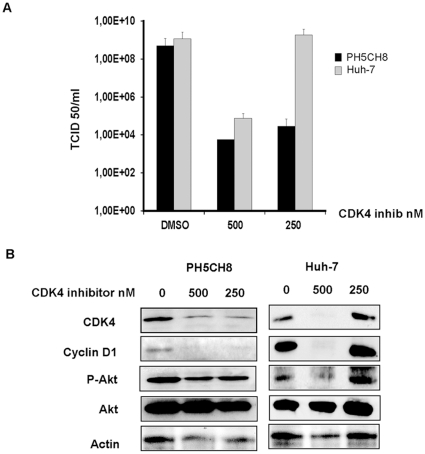
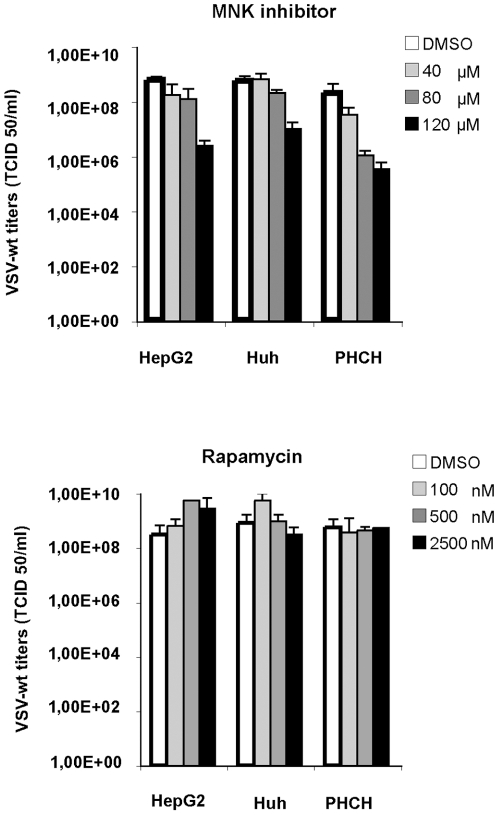
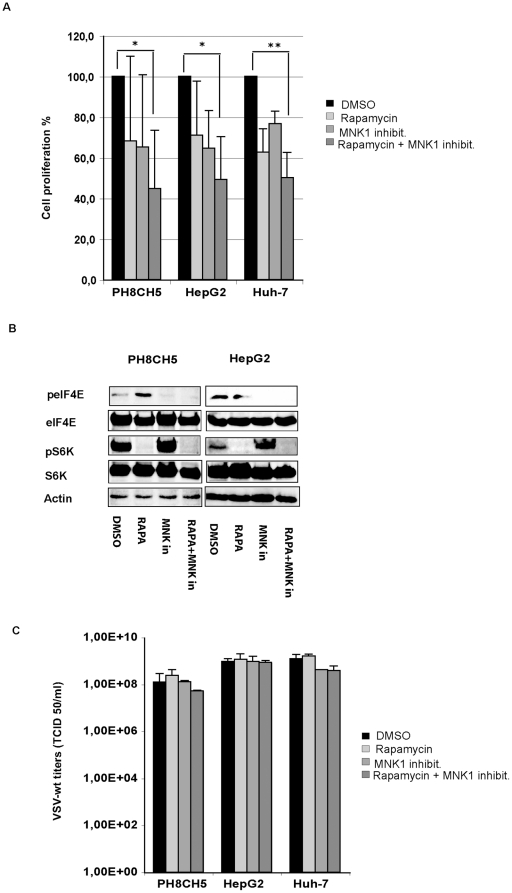
The intrinsic oncolytic specificity of vesicular stomatitis virus (VSV) is currently being exploited to develop alternative therapeutic strategies for hepatocellular carcinoma (HCC). Identifying key regulators in diverse transduction pathways that define VSV oncolysis in cancer cells represents a fundamental prerequisite to engineering more effective oncolytic viral vectors and adjusting combination therapies. After having identified defects in the signalling cascade of type I interferon induction, responsible for attenuated antiviral responses in human HCC cell lines, we have now investigated the role of cell proliferation and translation initiation. Cell cycle progression and translation initiation factors eIF4E and eIF2Bepsilon have been recently identified as key regulators of VSV permissiveness in T-lymphocytes and immortalized mouse embryonic fibroblasts, respectively. Here, we show that in HCC, decrease of cell proliferation by cell cycle inhibitors or siRNA-mediated reduction of G(1) cyclin-dependent kinase activities (CDK4) or cyclin D1 protein expression, do not significantly alter viral growth. Additionally, we demonstrate that translation initiation factors eIF4E and eIF2Bepsilon are negligible in sustaining VSV replication in HCC. Taken together, these results indicate that cellular proliferation and the initiation phase of cellular protein synthesis are not essential for successful VSV oncolysis of HCC. Moreover, our observations indicate the importance of cell-type specificity for VSV oncolysis, an important aspect to be considered in virotherapy applications in the future.
Conflict of interest statement
Figures








References
-
- Dancey JE. Therapeutic targets: MTOR and related pathways. Cancer Biol Ther. 2006;5:1065–1073. - PubMed
-
- Hallenbeck PL, Chang YN, Hay C, Golightly D, Stewart D, et al. A novel tumor-specific replication-restricted adenoviral vector for gene therapy of hepatocellular carcinoma. Hum Gene Ther. 1999;10:1721–1733. - PubMed
-
- Ebert O, Shinozaki K, Huang TG, Savontaus MJ, Garcia-Sastre A, et al. Oncolytic vesicular stomatitis virus for treatment of orthotopic hepatocellular carcinoma in immune-competent rats. Cancer Res. 2003;63:3605–3611. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
