

Disrupting circadian homeostasis of sympathetic signaling promotes tumor development in mice
- PMID: 20539819
- PMCID: PMC2881876
- DOI: 10.1371/journal.pone.0010995
Disrupting circadian homeostasis of sympathetic signaling promotes tumor development in mice
Abstract
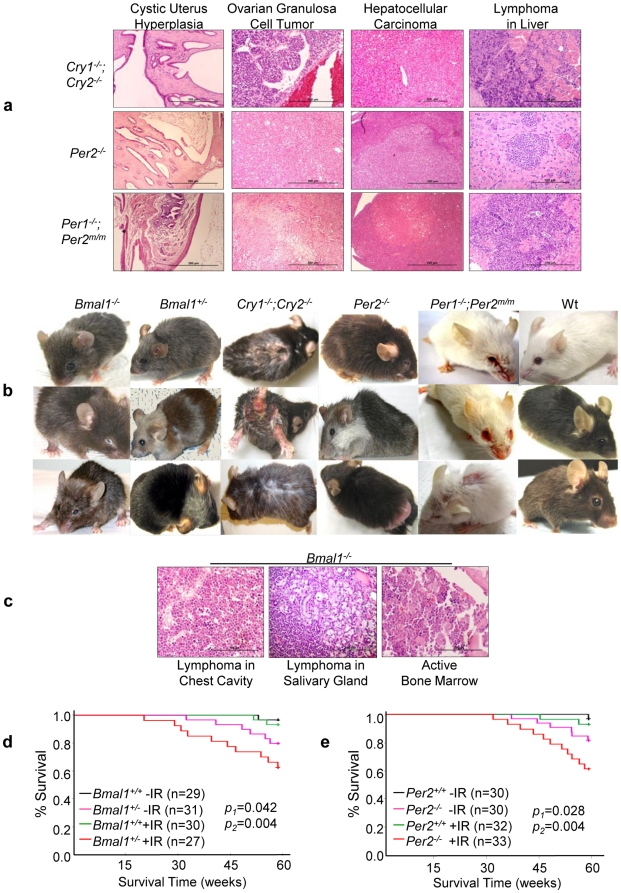
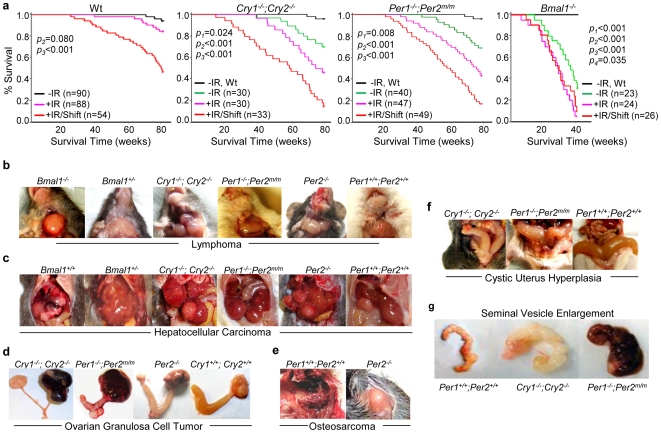
Background: Cell proliferation in all rapidly renewing mammalian tissues follows a circadian rhythm that is often disrupted in advanced-stage tumors. Epidemiologic studies have revealed a clear link between disruption of circadian rhythms and cancer development in humans. Mice lacking the circadian genes Period1 and 2 (Per) or Cryptochrome1 and 2 (Cry) are deficient in cell cycle regulation and Per2 mutant mice are cancer-prone. However, it remains unclear how circadian rhythm in cell proliferation is generated in vivo and why disruption of circadian rhythm may lead to tumorigenesis.
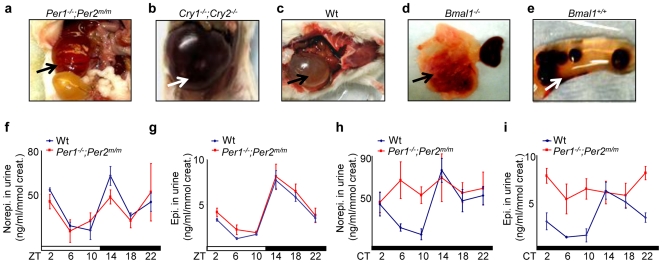
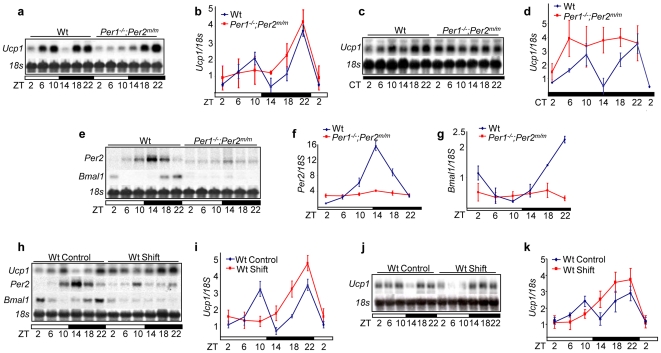
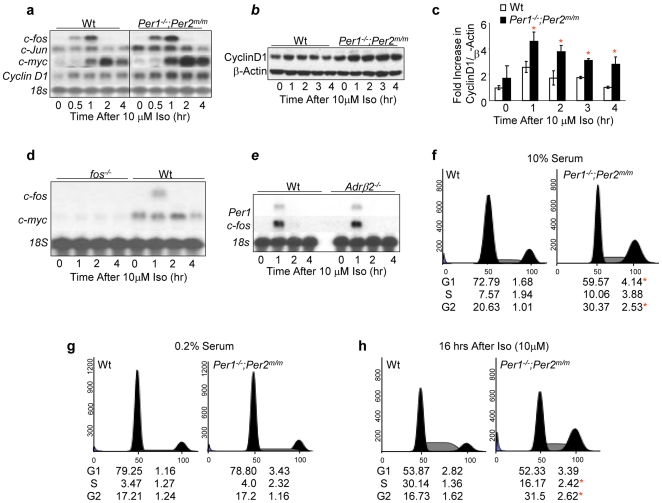
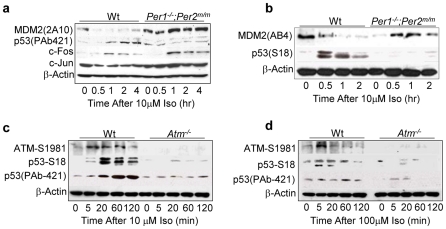
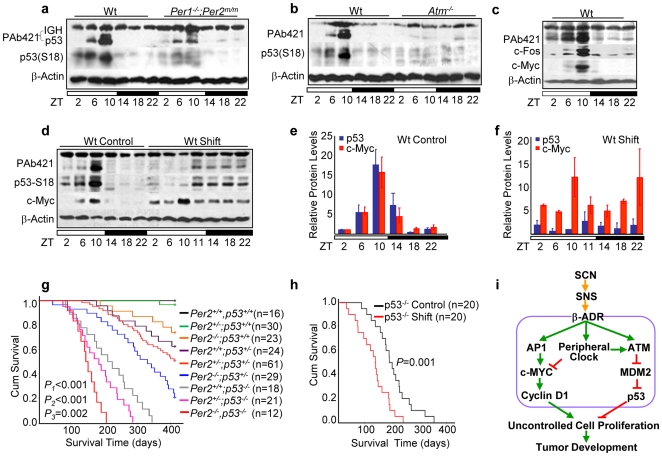
Methodology/principal findings: Mice lacking Per1 and 2, Cry1 and 2, or one copy of Bmal1, all show increased spontaneous and radiation-induced tumor development. The neoplastic growth of Per-mutant somatic cells is not controlled cell-autonomously but is dependent upon extracellular mitogenic signals. Among the circadian output pathways, the rhythmic sympathetic signaling plays a key role in the central-peripheral timing mechanism that simultaneously activates the cell cycle clock via AP1-controlled Myc induction and p53 via peripheral clock-controlled ATM activation. Jet-lag promptly desynchronizes the central clock-SNS-peripheral clock axis, abolishes the peripheral clock-dependent ATM activation, and activates myc oncogenic potential, leading to tumor development in the same organ systems in wild-type and circadian gene-mutant mice.
Conclusions/significance: Tumor suppression in vivo is a clock-controlled physiological function. The central circadian clock paces extracellular mitogenic signals that drive peripheral clock-controlled expression of key cell cycle and tumor suppressor genes to generate a circadian rhythm in cell proliferation. Frequent disruption of circadian rhythm is an important tumor promoting factor.
Conflict of interest statement
Figures
References
-
- Hamilton T. Influence of environmental light and melatonin upon mammary tumour induction. Br J Surg. 1969;56:764–766. - PubMed
-
- Aubert C, Janiaud P, Lecalvez J. Effect of pinealectomy and melatonin on mammary tumor growth in Sprague-Dawley rats under different conditions of lighting. J Neural Transm. 1980;47:121–130. - PubMed
-
- Shah PN, Mhatre MC, Kothari LS. Effect of melatonin on mammary carcinogenesis in intact and pinealectomized rats in varying photoperiods. Cancer Res. 1984;44:3403–3407. - PubMed
-
- van den Heiligenberg S, Depres-Brummer P, Barbason H, Claustrat B, Reynes M, et al. The tumor promoting effect of constant light exposure on diethylnitrosamine-induced hepatocarcinogenesis in rats. Life Sci. 1999;64:2523–2534. - PubMed
-
- Mhatre MC, Shah PN, Juneja HS. Effect of varying photoperiods on mammary morphology, DNA synthesis, and hormone profile in female rats. J Natl Cancer Inst. 1984;72:1411–1416. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
