

Influence of centriole number on mitotic spindle length and symmetry
- PMID: 20540087
- PMCID: PMC2923695
- DOI: 10.1002/cm.20462
Influence of centriole number on mitotic spindle length and symmetry
Abstract
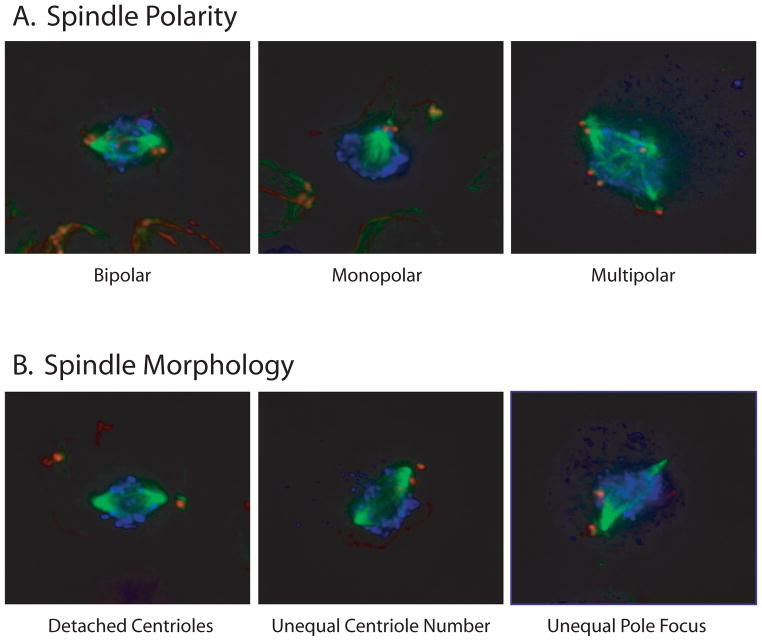
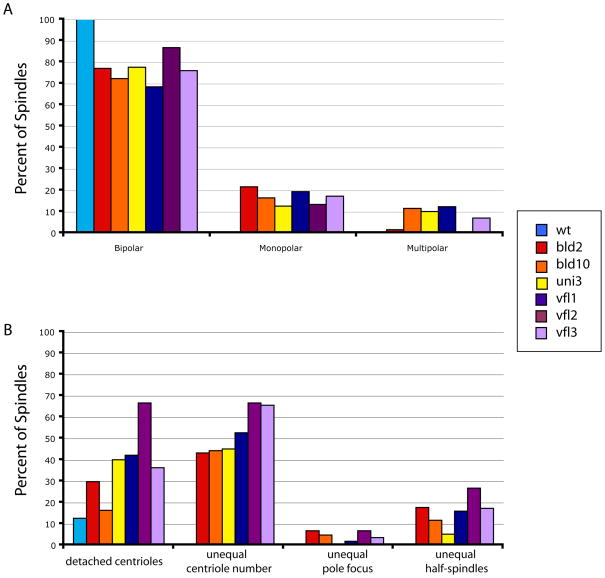
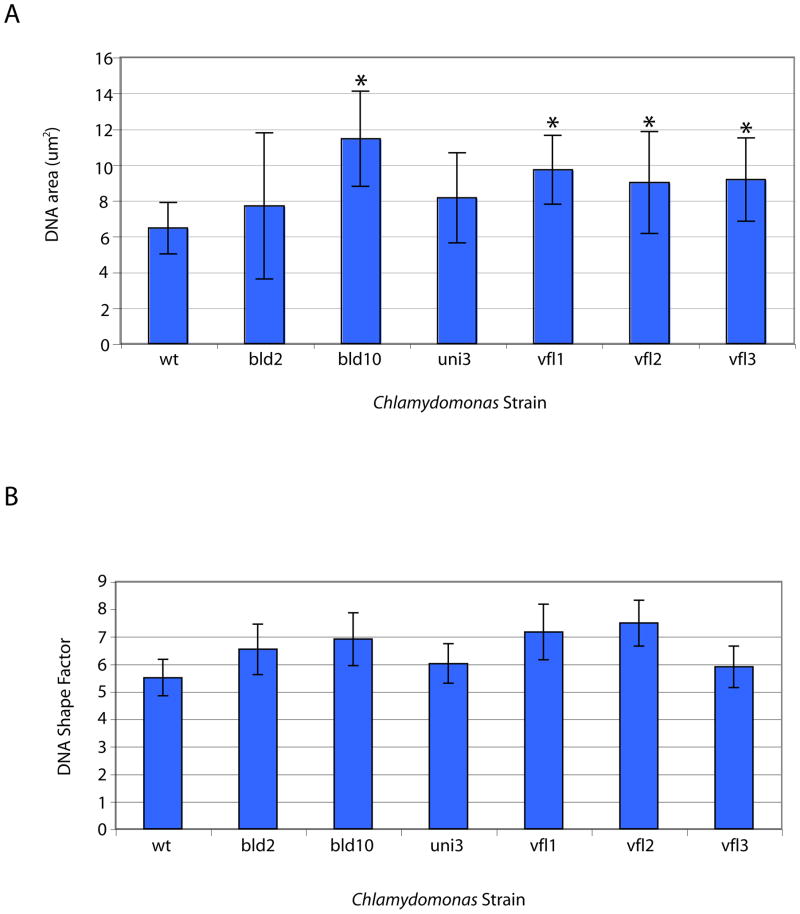
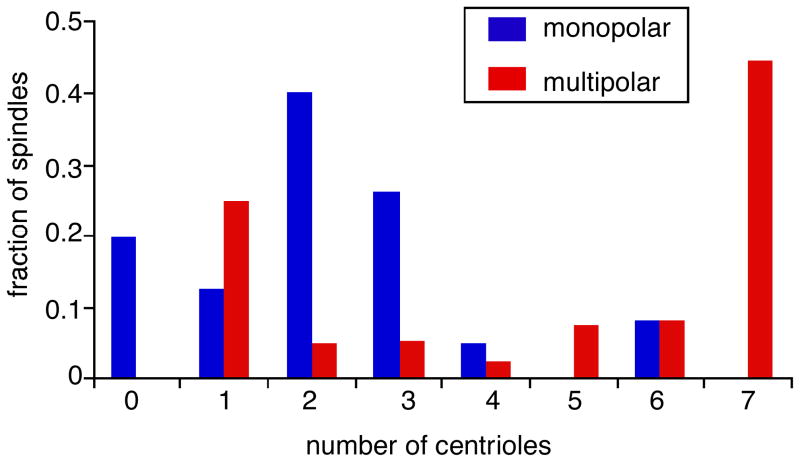
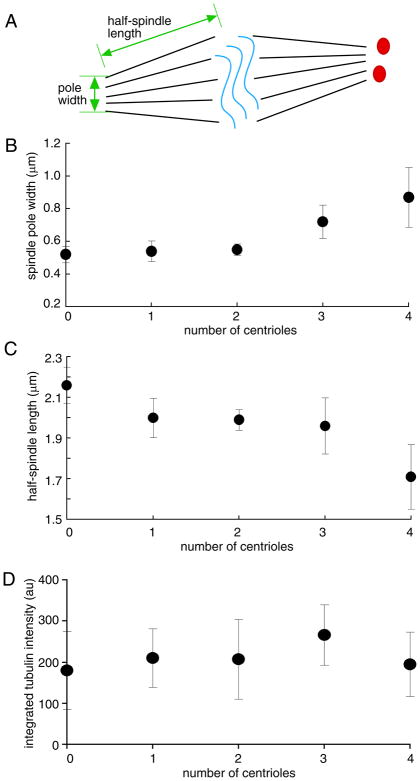
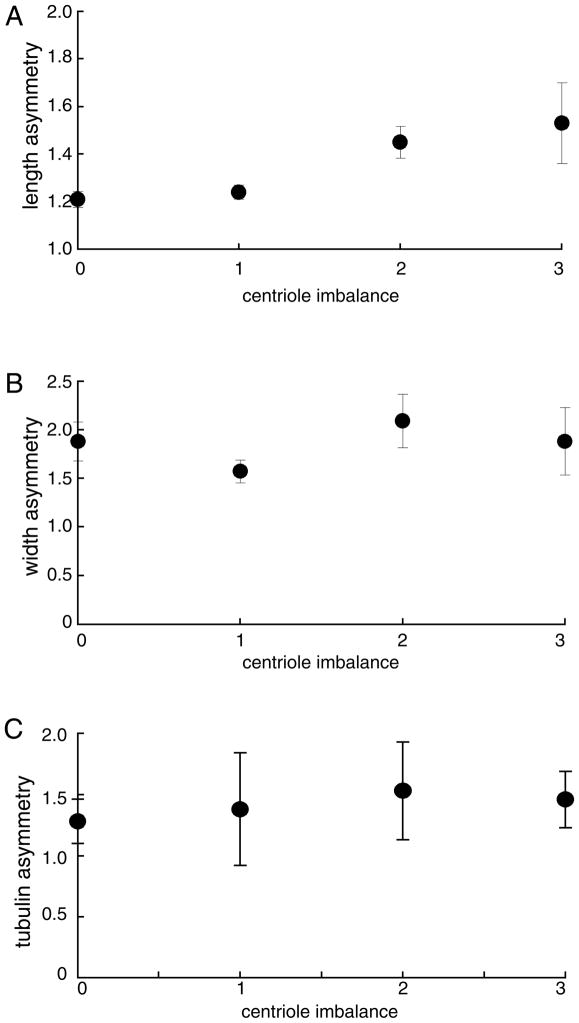
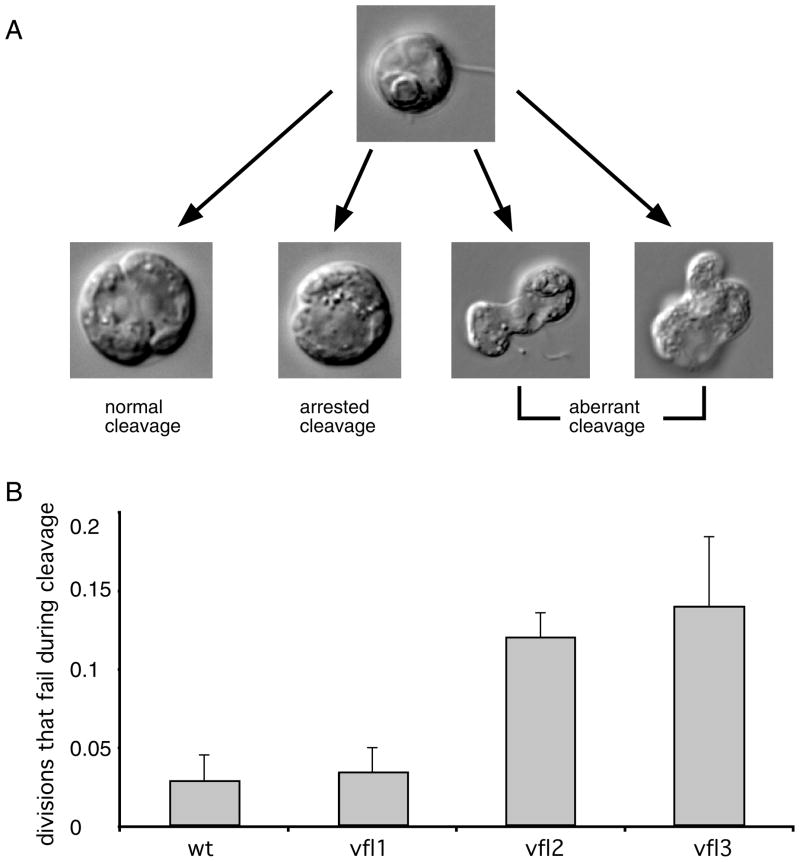
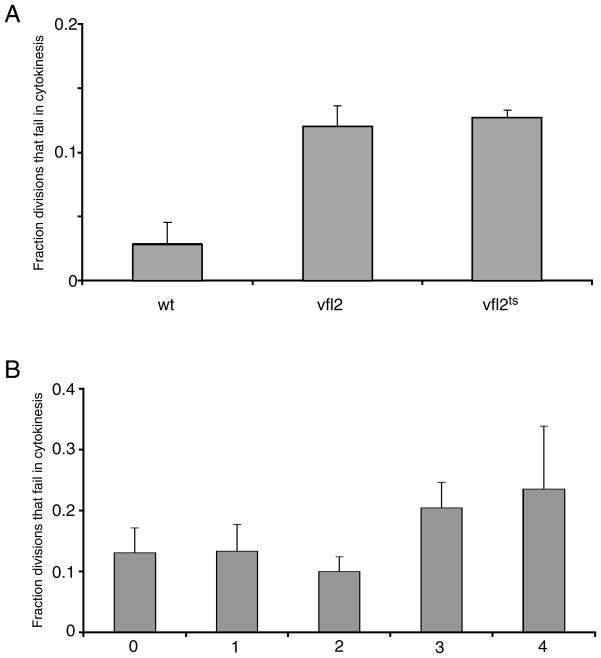
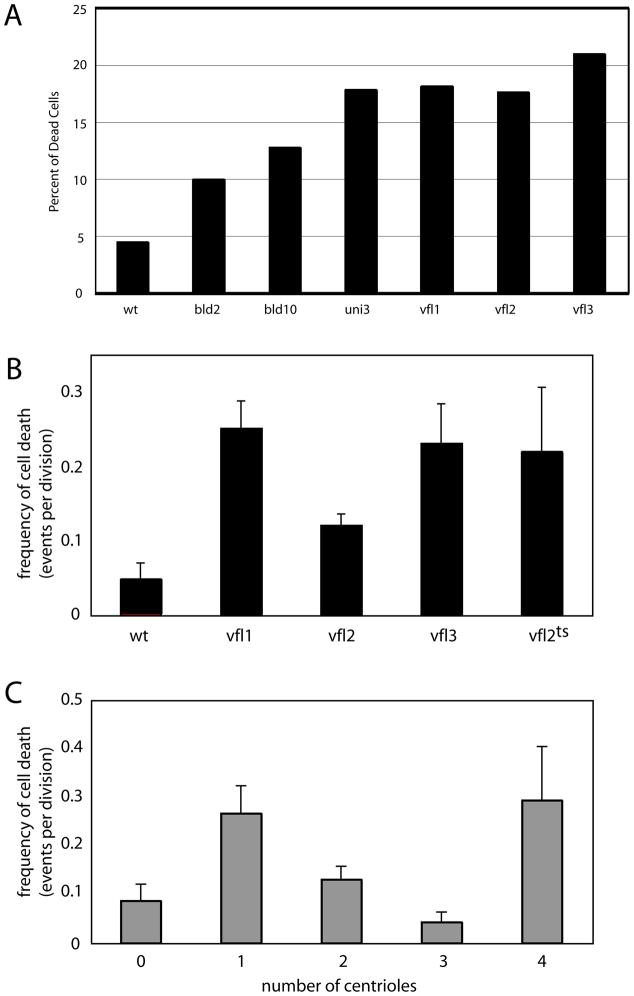
The functional role of centrioles or basal bodies in mitotic spindle assembly and function is currently unclear. Although supernumerary centrioles have been associated with multipolar spindles in cancer cells, suggesting centriole number might dictate spindle polarity, bipolar spindles are able to assemble in the complete absence of centrioles, suggesting a level of centriole-independence in the spindle assembly pathway. In this report we perturb centriole number using mutations in Chlamydomonas reinhardtii, and measure the response of the mitotic spindle to these perturbations in centriole number. Although altered centriole number increased the frequency of monopolar and multipolar spindles, the majority of spindles remained bipolar regardless of the centriole number. But even when spindles were bipolar, abnormal centriole numbers led to asymmetries in tubulin distribution, half-spindle length and spindle pole focus. Half spindle length correlated directly with number of centrioles at a pole, such that an imbalance in centriole number between the two poles of a bipolar spindle correlated with increased asymmetry between half spindle lengths. These results are consistent with centrioles playing an active role in regulating mitotic spindle length. Mutants with centriole number alteration also show increased cytokinesis defects, but these do not correlate with centriole number in the dividing cell and may therefore reflect downstream consequences of defects in preceding cell divisions.
2010 Wiley-Liss, Inc.
Figures
Similar articles
-
ASQ2 encodes a TBCC-like protein required for mother-daughter centriole linkage and mitotic spindle orientation.Curr Biol. 2009 Jul 28;19(14):1238-43. doi: 10.1016/j.cub.2009.05.071. Curr Biol. 2009. PMID: 19631545 Free PMC article.
-
Centriole number and the reproductive capacity of spindle poles.J Cell Biol. 1985 Mar;100(3):887-96. doi: 10.1083/jcb.100.3.887. J Cell Biol. 1985. PMID: 3972899 Free PMC article.
-
Centrosome-centriole abnormalities are markers for abnormal cell divisions and cancer in the transgenic adenocarcinoma mouse prostate (TRAMP) model.Biol Cell. 2000 Aug;92(5):331-40. doi: 10.1016/s0248-4900(00)01079-0. Biol Cell. 2000. PMID: 11071042
-
Cell division: The renaissance of the centriole.Curr Biol. 1999 Mar 25;9(6):R218-20. doi: 10.1016/s0960-9822(99)80133-x. Curr Biol. 1999. PMID: 10209087 Review.
-
Elucidation of basal body and centriole functions in Chlamydomonas reinhardtii.Traffic. 2003 Jul;4(7):443-51. doi: 10.1034/j.1600-0854.2003.00104.x. Traffic. 2003. PMID: 12795689 Review.
Cited by
-
Mitotic spindle formation in Triparma laevis NIES-2565(Parmales, Heterokontophyta).Protoplasma. 2017 Jan;254(1):461-471. doi: 10.1007/s00709-016-0967-x. Epub 2016 Apr 5. Protoplasma. 2017. PMID: 27048177
-
Scaling of Subcellular Structures.Annu Rev Cell Dev Biol. 2020 Oct 6;36:219-236. doi: 10.1146/annurev-cellbio-020520-113246. Epub 2020 Jun 30. Annu Rev Cell Dev Biol. 2020. PMID: 32603615 Free PMC article. Review.
-
Filaments and fingers: Novel structural aspects of the single septin from Chlamydomonas reinhardtii.J Biol Chem. 2017 Jun 30;292(26):10899-10911. doi: 10.1074/jbc.M116.762229. Epub 2017 May 5. J Biol Chem. 2017. PMID: 28476887 Free PMC article.
-
Centrosomes in spindle organization and chromosome segregation: a mechanistic view.Chromosome Res. 2016 Jan;24(1):19-34. doi: 10.1007/s10577-015-9508-2. Chromosome Res. 2016. PMID: 26643311 Review.
-
CCDC61/VFL3 Is a Paralog of SAS6 and Promotes Ciliary Functions.Structure. 2020 Jun 2;28(6):674-689.e11. doi: 10.1016/j.str.2020.04.010. Epub 2020 May 5. Structure. 2020. PMID: 32375023 Free PMC article.
References
-
- Abal M, Keryer G, Bornens M. Centrioles resist forces applied on centrosomes during G2/M transition. Biol Cell. 2005;97:425–34. - PubMed
-
- Basto R, Lau J, Vinogradova T, Gardiol A, Woods CG, Khodjakov A, Raff JW. Flies without centrioles. Cell. 2006;125:1375–86. - PubMed
-
- Boveri T. Zur Frage der Entstehung Maligner Tumoren. Jena, Germany: Fischer Verlag; 1914. (English translation by M. Boveri reprinted as The Origin of Malignant Tumors, The Williams and Wilkins Co., Baltimore, 1929.)
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
