

Sinoatrial tissue of crucian carp heart has only negative contractile responses to autonomic agonists
- PMID: 20540719
- PMCID: PMC2894799
- DOI: 10.1186/1472-6793-10-10
Sinoatrial tissue of crucian carp heart has only negative contractile responses to autonomic agonists
Abstract
Background: In the anoxia-tolerant crucian carp (Carassius carassius) cardiac activity varies according to the seasons. To clarify the role of autonomic nervous control in modulation of cardiac activity, responses of atrial contraction and heart rate (HR) to carbacholine (CCh) and isoprenaline (Iso) were determined in fish acclimatized to winter (4 degrees C, cold-acclimated, CA) and summer (18 degrees C, warm-acclimated, WA) temperatures.
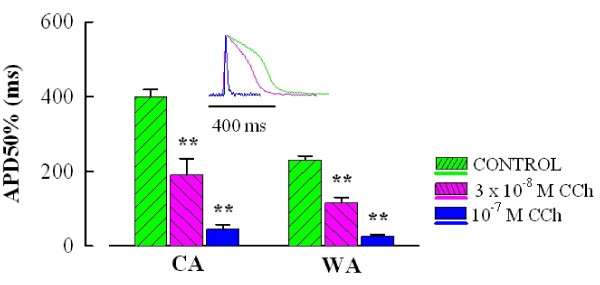
Results: Inhibitory action of CCh was much stronger on atrial contractility than HR. CCh reduced force of atrial contraction at an order of magnitude lower concentrations (EC50 2.75-3.5.10-8 M) in comparison to its depressive effect on HR (EC50 1.23-2.02.10-7 M) (P < 0.05) without differences between winter and summer acclimatized fish. Inhibition of nitric oxide synthase with 100 microM L-NMMA did not change the response of the sinoatrial tissue to CCh. Reduction of atrial force was associated with a strong shortening of action potential (AP) duration to approximately 50% (48 +/- 10 and 50 +/- 6% for CA and WA fish, respectively) and 11% (11 +/- 3 and 11 +/- 2% for CA and WA fish, respectively) of the control value at 3.10-8 M and 10-7 M CCh, respectively (P < 0.05). In atrial myocytes, CCh induced an inwardly rectifying K+ current, IK,CCh, with an EC50 value of 3-4.5.10-7 M and inhibited Ca2+ current (ICa) by 28 +/- 8% and 51 +/- 6% at 10-7 M and 10-6 M, respectively. These currents can explain the shortening of AP. Iso did not elicit any responses in crucian carp sinoatrial preparations nor did it have any effect on atrial ICa, probably due to the saturation of the beta-adrenergic cascade in the basal state.
Conclusion: In the crucian carp, HR and force of atrial contraction show cardio-depressive responses to the cholinergic agonist, but do not have any responses to the beta-adrenergic agonist. The scope of inhibitory regulation by CCh is increased by the high basal tone of the adenylate cyclase-cAMP cascade. Higher concentrations of CCh were required to induce IK,CCh and inhibit ICa than was needed for CCh's negative inotropic effect on atrial muscle suggesting that neither IK,CCh nor ICa alone can mediate CCh's actions but they might synergistically reduce AP duration and atrial force production. Autonomic responses were similar in CA winter fish and WA summer fish indicating that cardiac sensitivity to external modulation by the autonomic nervous system is not involved in seasonal acclimatization of the crucian carp heart to cold and anoxic winter conditions.
Figures







Similar articles
-
Electrophysiological differences in cholinergic signaling between the hearts of summer and winter frogs (Rana temporaria).J Comp Physiol B. 2018 Jul;188(4):649-656. doi: 10.1007/s00360-018-1147-4. Epub 2018 Feb 10. J Comp Physiol B. 2018. PMID: 29429060
-
Seasonal changes of cholinergic response in the atrium of Arctic navaga cod (Eleginus navaga).J Comp Physiol B. 2017 Feb;187(2):329-338. doi: 10.1007/s00360-016-1032-y. Epub 2016 Sep 26. J Comp Physiol B. 2017. PMID: 27672043
-
A cellular mechanism for nitric oxide-mediated cholinergic control of mammalian heart rate.J Gen Physiol. 1995 Jul;106(1):45-65. doi: 10.1085/jgp.106.1.45. J Gen Physiol. 1995. PMID: 7494138 Free PMC article.
-
Nitric oxide regulation of atrioventricular node excitability.Can J Cardiol. 1997 Dec;13(12):1191-201. Can J Cardiol. 1997. PMID: 9444302 Review.
-
The force-frequency relationship: insights from mathematical modeling.Adv Physiol Educ. 2013 Mar;37(1):28-34. doi: 10.1152/advan.00072.2011. Adv Physiol Educ. 2013. PMID: 23471245 Free PMC article. Review.
Cited by
-
Electrophysiological differences in cholinergic signaling between the hearts of summer and winter frogs (Rana temporaria).J Comp Physiol B. 2018 Jul;188(4):649-656. doi: 10.1007/s00360-018-1147-4. Epub 2018 Feb 10. J Comp Physiol B. 2018. PMID: 29429060
-
Seasonal changes of cholinergic response in the atrium of Arctic navaga cod (Eleginus navaga).J Comp Physiol B. 2017 Feb;187(2):329-338. doi: 10.1007/s00360-016-1032-y. Epub 2016 Sep 26. J Comp Physiol B. 2017. PMID: 27672043
-
Contractile performance of the Alaska blackfish (Dallia pectoralis) ventricle: Assessment of the effects of temperature, pacing frequency, the role of the sarcoplasmic reticulum in contraction and adrenergic stimulation.Comp Biochem Physiol A Mol Integr Physiol. 2019 Dec;238:110564. doi: 10.1016/j.cbpa.2019.110564. Epub 2019 Sep 4. Comp Biochem Physiol A Mol Integr Physiol. 2019. PMID: 31493554 Free PMC article.
-
Autonomic Regulation of the Goldfish Intact Heart.Front Physiol. 2022 Feb 9;13:793305. doi: 10.3389/fphys.2022.793305. eCollection 2022. Front Physiol. 2022. PMID: 35222073 Free PMC article.
-
The effective use of blebbistatin to study the action potential of cardiac pacemaker cells of zebrafish (Danio rerio) during incremental warming.Curr Res Physiol. 2022 Jan 17;5:48-54. doi: 10.1016/j.crphys.2022.01.002. eCollection 2022. Curr Res Physiol. 2022. PMID: 35128467 Free PMC article.
References
-
- Holopainen IJ, Tonn WM, Paszkowski CA. Tales of two fish: the dichotomous biology of crucian carp (Carassius carassius (L.)) in northern Europe. Ann Zool Fenn. 1997;28:1–22.
-
- Piironen J, Holopainen IJ. A note on seasonality in anoxia tolerance of crucian carp (Carassius carassius L.) in the laboratory. Ann Zool Fenn. 1986;23:335–338.
-
- Vornanen M, Paajanen V. Seasonality of dihydropyridine receptor binding in the heart of an anoxia-tolerant vertebrate, the crucian carp (Carassius carassius L.) Am J Physiol. 2004;287:R1263–R1269. - PubMed
-
- Johnston IA, Bernard LM. Utilization of the ethanol pathway in carp following exposure to anoxia. J Exp Biol. 1983;104:73–78.
-
- Holopainen IJ, Hyvärinen H. Ecology and physiology of crucian carp (Carassius carassius (L.)) in small Finnish ponds with anoxic conditions in winter. Verh Internat Verein Limnol. 1985;22:2566–2570.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
