

Roles of CUP-5, the Caenorhabditis elegans orthologue of human TRPML1, in lysosome and gut granule biogenesis
- PMID: 20540742
- PMCID: PMC2891664
- DOI: 10.1186/1471-2121-11-40
Roles of CUP-5, the Caenorhabditis elegans orthologue of human TRPML1, in lysosome and gut granule biogenesis
Abstract
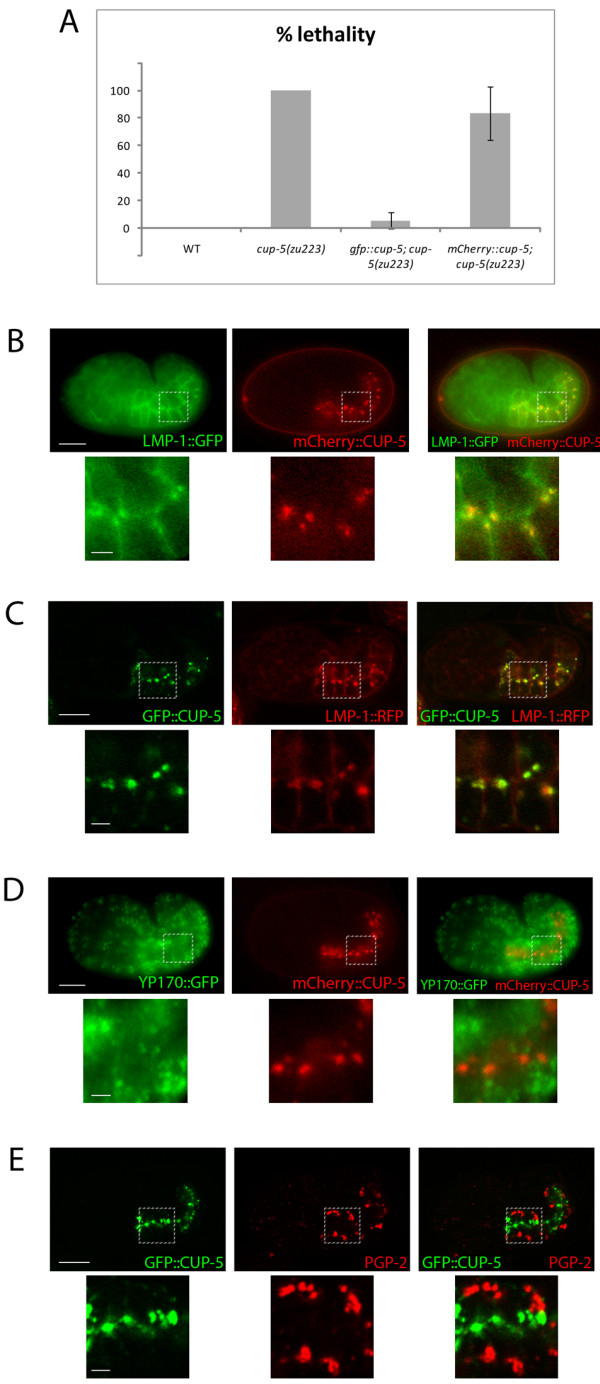
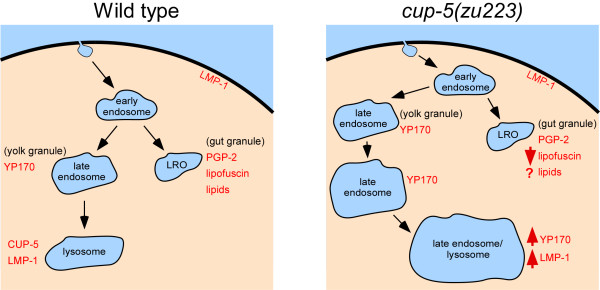
Background: CUP-5 is a Transient Receptor Potential protein in C. elegans that is the orthologue of mammalian TRPML1. Loss of TRPML1 results in the lysosomal storage disorder Mucolipidosis type IV. Loss of CUP-5 results in embryonic lethality and the accumulation of enlarged yolk granules in developing intestinal cells. The embryonic lethality of cup-5 mutants is rescued by mutations in mrp-4, which is required for gut granule differentiation. Gut granules are intestine-specific lysosome-related organelles that accumulate birefringent material. This link between CUP-5 and gut granules led us to determine the roles of CUP-5 in lysosome and gut granule biogenesis in developing intestinal cells.
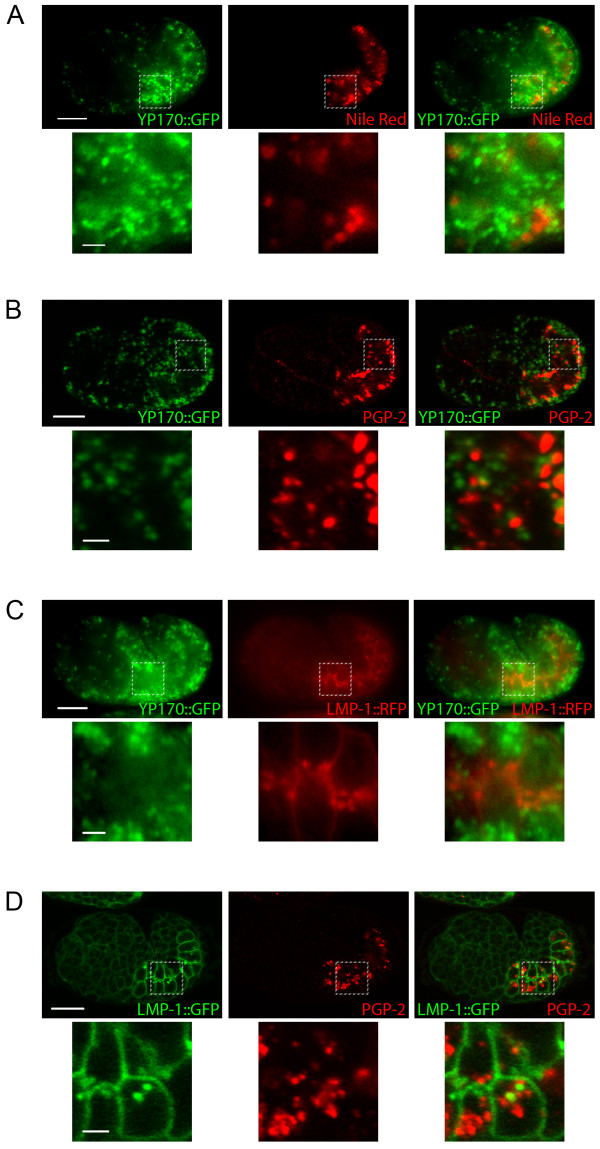
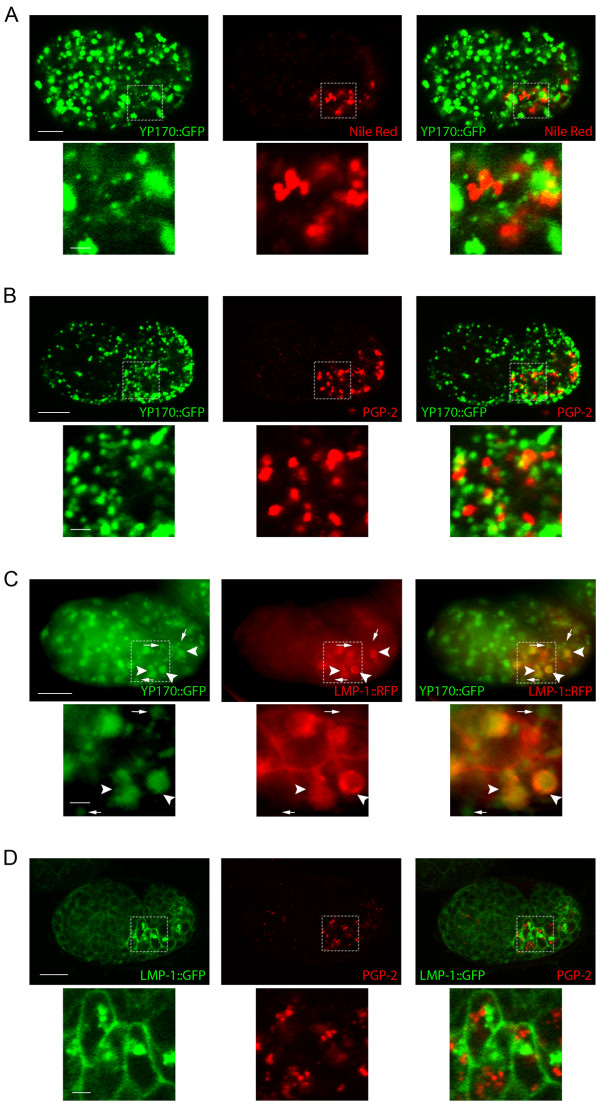
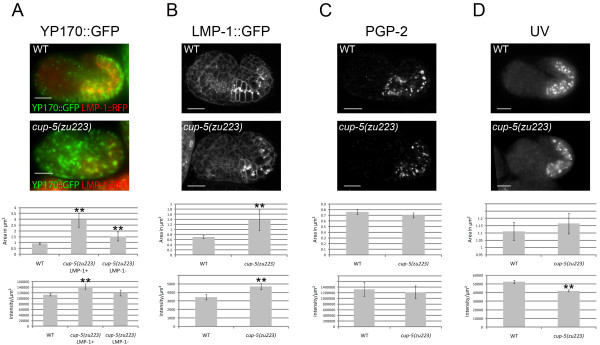
Results: We show that CUP-5 protein localizes to lysosomes, but not to gut granules, in developing intestinal cells. Loss of CUP-5 results in defects in endo-lysosomal transport in developing intestinal cells of C. elegans embryos. This ultimately leads to the appearance of enlarged terminal vacuoles that show defective lysosomal degradation and that have lysosomal and endosomal markers. In contrast, gut granule biogenesis is normal in the absence of CUP-5. Furthermore, loss of CUP-5 does not result in inappropriate fusion or mixing of content between lysosomes and gut granules.
Conclusions: Using an in vivo model of MLIV, we show that there is a defect in lysosomal transport/biogenesis that is earlier than the presumed function of TRPML1 in terminal lysosomes. Our results indicate that CUP-5 is required for the biogenesis of lysosomes but not of gut granules. Thus, cellular phenotypes in Mucolipidosis type IV are likely not due to defects in lysosome-related organelle biogenesis, but due to progressive defects in lysosomal transport that lead to severe lysosomal dysfunction.
Figures
Similar articles
-
Suppression of the cup-5 mucolipidosis type IV-related lysosomal dysfunction by the inactivation of an ABC transporter in C. elegans.Development. 2006 Oct;133(19):3939-48. doi: 10.1242/dev.02575. Epub 2006 Aug 30. Development. 2006. PMID: 16943270
-
ESCRT-Dependent Cell Death in a Caenorhabditis elegans Model of the Lysosomal Storage Disorder Mucolipidosis Type IV.Genetics. 2016 Feb;202(2):619-38. doi: 10.1534/genetics.115.182485. Epub 2015 Nov 23. Genetics. 2016. PMID: 26596346 Free PMC article.
-
Basis of lethality in C. elegans lacking CUP-5, the Mucolipidosis Type IV orthologue.Dev Biol. 2006 May 15;293(2):382-91. doi: 10.1016/j.ydbio.2006.02.008. Dev Biol. 2006. PMID: 16530747
-
Caenorhabditis elegans as a model for lysosomal storage disorders.Biochim Biophys Acta. 2008 Jul-Aug;1782(7-8):433-46. doi: 10.1016/j.bbadis.2008.04.003. Epub 2008 May 1. Biochim Biophys Acta. 2008. PMID: 18501720 Review.
-
CUPpling calcium to lysosomal biogenesis.Trends Cell Biol. 2004 Sep;14(9):471-3. doi: 10.1016/j.tcb.2004.07.010. Trends Cell Biol. 2004. PMID: 15350973 Review.
Cited by
-
A zinc metal complex as an NIR emissive probe for real-time dynamics and in vivo embryogenic evolution of lysosomes using super-resolution microscopy.Chem Sci. 2024 Sep 5;15(38):15659-69. doi: 10.1039/d4sc04638b. Online ahead of print. Chem Sci. 2024. PMID: 39246364 Free PMC article.
-
ADF/cofilin promotes invadopodial membrane recycling during cell invasion in vivo.J Cell Biol. 2014 Mar 31;204(7):1209-18. doi: 10.1083/jcb.201312098. Epub 2014 Mar 24. J Cell Biol. 2014. PMID: 24662568 Free PMC article.
-
Caenorhabditis elegans HOPS and CCZ-1 mediate trafficking to lysosome-related organelles independently of RAB-7 and SAND-1.Mol Biol Cell. 2014 Apr;25(7):1073-96. doi: 10.1091/mbc.E13-09-0521. Epub 2014 Feb 5. Mol Biol Cell. 2014. PMID: 24501423 Free PMC article.
-
Function and regulation of the Caenorhabditis elegans Rab32 family member GLO-1 in lysosome-related organelle biogenesis.PLoS Genet. 2018 Nov 12;14(11):e1007772. doi: 10.1371/journal.pgen.1007772. eCollection 2018 Nov. PLoS Genet. 2018. PMID: 30419011 Free PMC article.
-
A pro-cathepsin L mutant is a luminal substrate for endoplasmic-reticulum-associated degradation in C. elegans.PLoS One. 2012;7(7):e40145. doi: 10.1371/journal.pone.0040145. Epub 2012 Jul 2. PLoS One. 2012. PMID: 22768338 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
