

Evolution in action: N and C termini of subunits in related T = 4 viruses exchange roles as molecular switches
- PMID: 20541507
- PMCID: PMC2913159
- DOI: 10.1016/j.str.2010.03.010
Evolution in action: N and C termini of subunits in related T = 4 viruses exchange roles as molecular switches
Abstract
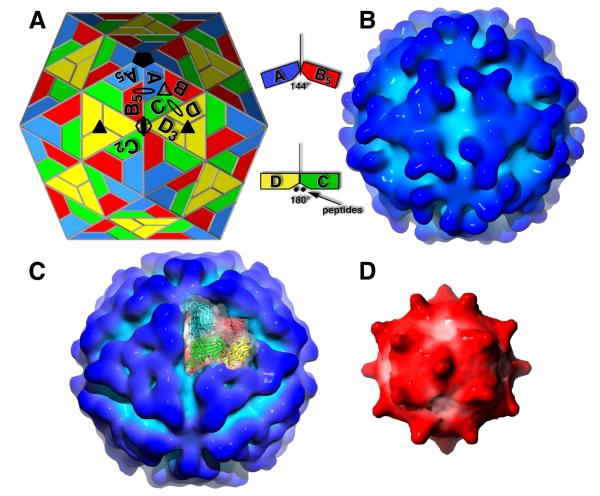
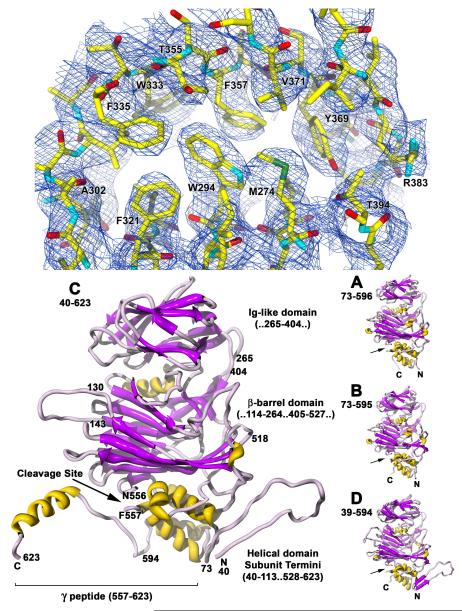
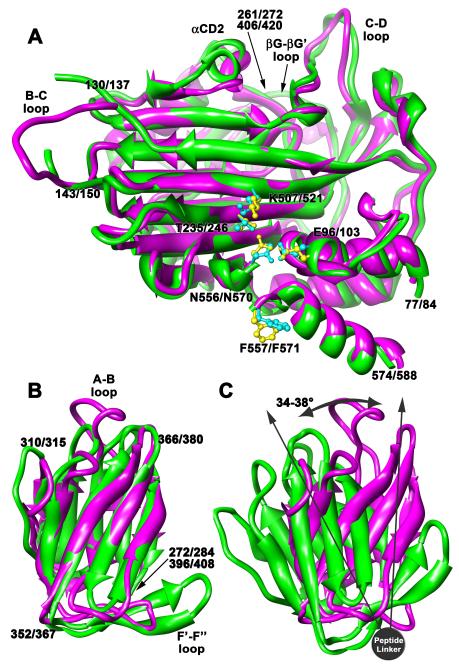
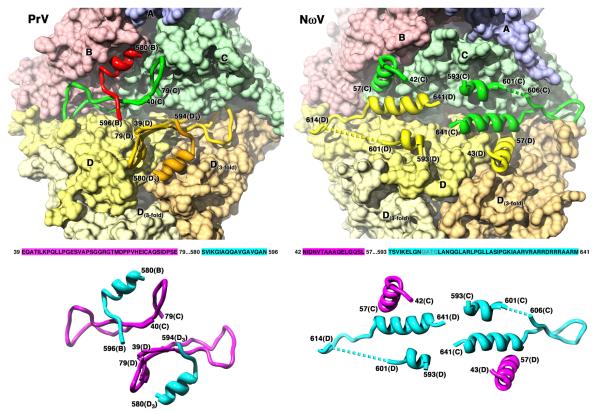
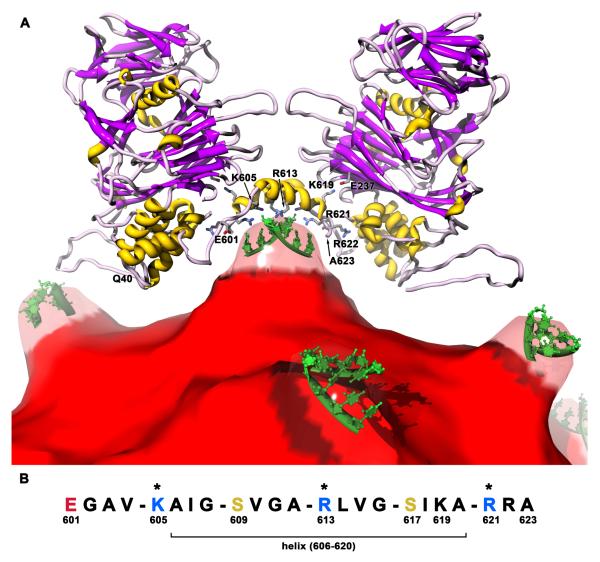
The T = 4 tetravirus and T = 3 nodavirus capsid proteins undergo closely similar autoproteolysis to produce the N-terminal beta and C-terminal, lipophilic gamma polypeptides. The gamma peptides and the N termini of beta also act as molecular switches that determine their quasi equivalent capsid structures. The crystal structure of Providence virus (PrV), only the second of a tetravirus (the first was NomegaV), reveals conserved folds and cleavage sites, but the protein termini have completely different structures and the opposite functions of those in NomegaV. N termini of beta form the molecular switch in PrV, whereas gamma peptides play this role in NomegaV. PrV gamma peptides instead interact with packaged RNA at the particle two-folds by using a repeating sequence pattern found in only four other RNA- or membrane-binding proteins. The disposition of peptide termini in PrV is closely related to those in nodaviruses, suggesting that PrV may be closer to the primordial T = 4 particle than NomegaV.
Figures
Similar articles
-
Dissecting quasi-equivalence in nonenveloped viruses: membrane disruption is promoted by lytic peptides released from subunit pentamers, not hexamers.J Virol. 2012 Sep;86(18):9976-82. doi: 10.1128/JVI.01089-12. Epub 2012 Jul 3. J Virol. 2012. PMID: 22761380 Free PMC article.
-
The refined structure of Nudaurelia capensis omega virus reveals control elements for a T = 4 capsid maturation.Virology. 2004 Jan 5;318(1):192-203. doi: 10.1016/j.virol.2003.08.045. Virology. 2004. PMID: 14972547
-
The three-dimensional structure of ryegrass mottle virus at 2.9 A resolution.Virology. 2007 Dec 20;369(2):364-74. doi: 10.1016/j.virol.2007.07.028. Epub 2007 Sep 19. Virology. 2007. PMID: 17881031
-
The Basic Architecture of Viruses.Subcell Biochem. 2024;105:55-78. doi: 10.1007/978-3-031-65187-8_2. Subcell Biochem. 2024. PMID: 39738944 Review.
-
What does structure tell us about virus evolution?Curr Opin Struct Biol. 2005 Dec;15(6):655-63. doi: 10.1016/j.sbi.2005.10.012. Epub 2005 Nov 3. Curr Opin Struct Biol. 2005. PMID: 16271469 Review.
Cited by
-
Assembly and maturation of a T = 4 quasi-equivalent virus is guided by electrostatic and mechanical forces.Viruses. 2014 Aug 22;6(8):3348-62. doi: 10.3390/v6083348. Viruses. 2014. PMID: 25153346 Free PMC article. Review.
-
Nucleic acid packaging in viruses.Curr Opin Struct Biol. 2012 Feb;22(1):65-71. doi: 10.1016/j.sbi.2011.11.002. Epub 2012 Jan 23. Curr Opin Struct Biol. 2012. PMID: 22277169 Free PMC article. Review.
-
Dissecting quasi-equivalence in nonenveloped viruses: membrane disruption is promoted by lytic peptides released from subunit pentamers, not hexamers.J Virol. 2012 Sep;86(18):9976-82. doi: 10.1128/JVI.01089-12. Epub 2012 Jul 3. J Virol. 2012. PMID: 22761380 Free PMC article.
-
Molecular and biochemical characterization of the NS1 protein of non-cultured influenza B virus strains circulating in Singapore.Microb Genom. 2016 Aug 25;2(8):e000082. doi: 10.1099/mgen.0.000082. eCollection 2016 Aug. Microb Genom. 2016. PMID: 28348873 Free PMC article.
-
Structure of the T=13 capsid of infectious pancreatic necrosis virus (IPNV)-a salmonid birnavirus.J Virol. 2025 Feb 25;99(2):e0145424. doi: 10.1128/jvi.01454-24. Epub 2025 Jan 16. J Virol. 2025. PMID: 39817769 Free PMC article.
References
-
- Bothner B, Dong XF, Bibbs L, Johnson JE, Siuzdak G. Evidence of viral capsid dynamics using limited proteolysis and mass spectrometry. J Biol Chem. 1998;273:673–676. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
