

'And then there were three': highly efficient uptake of potassium by foliar trichomes of epiphytic bromeliads
- PMID: 20542886
- PMCID: PMC2924823
- DOI: 10.1093/aob/mcq120
'And then there were three': highly efficient uptake of potassium by foliar trichomes of epiphytic bromeliads
Abstract
Background and aims: Vascular epiphytes have to acquire nutrients from atmospheric wash out, stem-flow, canopy soils and trapped litter. Physiological studies on the adaptations to nutrient acquisition and plant utilization of nutrients have focused on phosphorus and nitrogen; potassium, as a third highly abundant nutrient element, has received minor attention. In the present study, potassium uptake kinetics by leaves, within-plant distribution and nutrient accumulation were analysed to gain an improved understanding of physiological adaptations to non-terrestrial nutrient supply of plants.
Methods: Radioactively labelled (86)RbCl was used as an analogue to study uptake kinetics of potassium absorbed from tanks of epiphytes, its plant distribution and the correlation between uptake efficiency and abundance of trichomes, functioning as uptake organs of leaves. Potassium in leaves was additionally analysed by atomic absorption spectroscopy to assess plant responses to potassium deficiency.
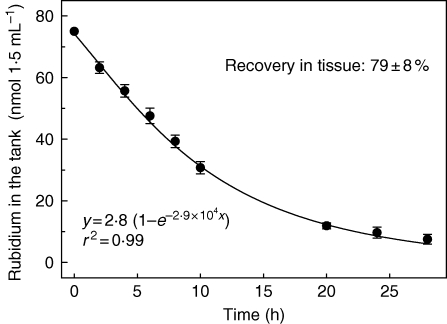
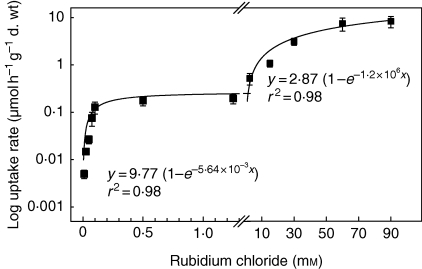
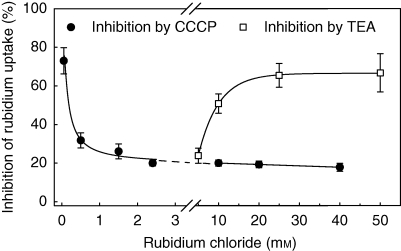
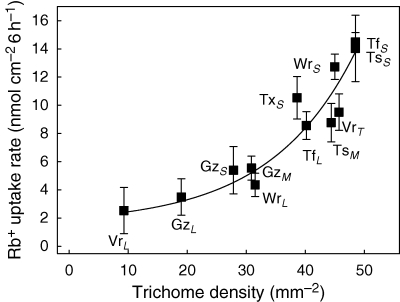
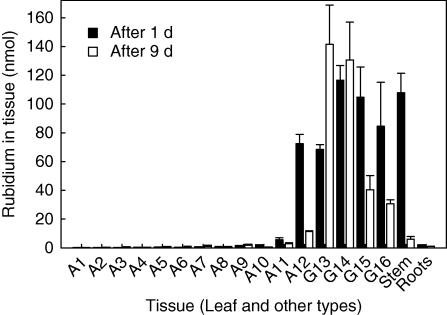
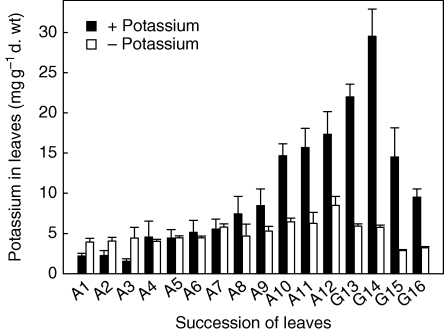
Key results: Labelled rubidium was taken up from tanks over a wide range of concentrations, 0.01-90 mm, which was achieved by two uptake systems. In four tank epiphytes, the high-affinity transporters had average K(m) values of 41.2 microm, and the low-affinity transporters average K(m) values of 44.8 mm. Further analysis in Vriesea splenriet showed that high-affinity uptake of rubidium was an ATP-dependent process, while low-affinity uptake was mediated by a K(+)-channel. The kinetic properties of both types of transporters are comparable with those of potassium transporters in roots of terrestrial plants. Specific differences in uptake velocities of epiphytes are correlated with the abundance of trichomes on their leaf surfaces. The main sinks for potassium were fully grown leaves. These leaves thus function as internal potassium sources, which allow growth to be maintained during periods of low external potassium availability.
Conclusions: Vascular epiphytes possess effective mechanisms to take up potassium from both highly diluted and highly concentrated solutions, enabling the plant to incorporate this nutrient element quickly and almost quantitatively from tank solutions. A surplus not needed for current metabolism is stored, i.e. plants show luxury consumption.
Figures
Similar articles
-
Highly efficient uptake of phosphorus in epiphytic bromeliads.Ann Bot. 2009 Feb;103(3):477-84. doi: 10.1093/aob/mcn231. Epub 2008 Nov 25. Ann Bot. 2009. PMID: 19033287 Free PMC article.
-
Microbial activities and foliar uptake of nitrogen in the epiphytic bromeliad Vriesea gigantea.New Phytol. 2007;175(2):311-320. doi: 10.1111/j.1469-8137.2007.02098.x. New Phytol. 2007. PMID: 17587379
-
Are vascular epiphytes nitrogen or phosphorus limited? A study of plant (15) N fractionation and foliar N : P stoichiometry with the tank bromeliad Vriesea sanguinolenta.New Phytol. 2011 Oct;192(2):462-70. doi: 10.1111/j.1469-8137.2011.03812.x. Epub 2011 Jul 6. New Phytol. 2011. PMID: 21729088
-
NPKS uptake, sensing, and signaling and miRNAs in plant nutrient stress.Protoplasma. 2016 May;253(3):767-786. doi: 10.1007/s00709-015-0845-y. Epub 2015 Jun 18. Protoplasma. 2016. PMID: 26085375 Review.
-
High-affinity potassium and sodium transport systems in plants.J Exp Bot. 2006;57(5):1149-60. doi: 10.1093/jxb/erj068. Epub 2006 Jan 31. J Exp Bot. 2006. PMID: 16449373 Review.
Cited by
-
Aerial roots of epiphytic orchids: the velamen radicum and its role in water and nutrient uptake.Oecologia. 2013 Mar;171(3):733-41. doi: 10.1007/s00442-012-2575-6. Epub 2013 Jan 6. Oecologia. 2013. PMID: 23292456
-
Effects of changes in leaf properties mediated by methyl jasmonate (MeJA) on foliar absorption of Zn, Mn and Fe.Ann Bot. 2017 Sep 1;120(3):405-415. doi: 10.1093/aob/mcx063. Ann Bot. 2017. PMID: 28641371 Free PMC article.
-
Absorption of foliar-applied Zn in sunflower (Helianthus annuus): importance of the cuticle, stomata and trichomes.Ann Bot. 2019 Jan 1;123(1):57-68. doi: 10.1093/aob/mcy135. Ann Bot. 2019. PMID: 30020418 Free PMC article.
-
Ecological stoichiometry of the epiphyte community in a subtropical forest canopy.Ecol Evol. 2019 Nov 28;9(24):14394-14406. doi: 10.1002/ece3.5875. eCollection 2019 Dec. Ecol Evol. 2019. PMID: 31938527 Free PMC article.
-
Absorption of foliar-applied Zn fertilizers by trichomes in soybean and tomato.J Exp Bot. 2018 Apr 27;69(10):2717-2729. doi: 10.1093/jxb/ery085. J Exp Bot. 2018. PMID: 29514247 Free PMC article.
References
-
- Adlassnig W, Steinhauser, Peroutka M, et al. Uptake of potassium, iron and manganese by carnivorous pitcher plants. Applied Radiation and Isotopes. 2009;67:2117–2122. - PubMed
-
- Benzing DH. Availability of exogenously supplied nitrogen to seedlings of Bromeliaceae. Bulletin of the Torrey Botanical Club. 1970;97:154–159.
-
- Benzing DH. Vascular epiphytes. General biology and related biota. Cambridge: Cambridge University Press; 1990.
-
- Benzing DH. Bromeliaceae. Profile of an adaptive radiation. Cambridge: Cambridge University Press; 2000.
-
- Benzing DH, Pridgeon AM. Foliar trichomes of Pleurothallidinae (Orchidaceae): functional significance. American Journal of Botany. 1983;70:173–180.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
