

An IL-2 paradox: blocking CD25 on T cells induces IL-2-driven activation of CD56(bright) NK cells
- PMID: 20543101
- PMCID: PMC3085179
- DOI: 10.4049/jimmunol.0902238
An IL-2 paradox: blocking CD25 on T cells induces IL-2-driven activation of CD56(bright) NK cells
Abstract
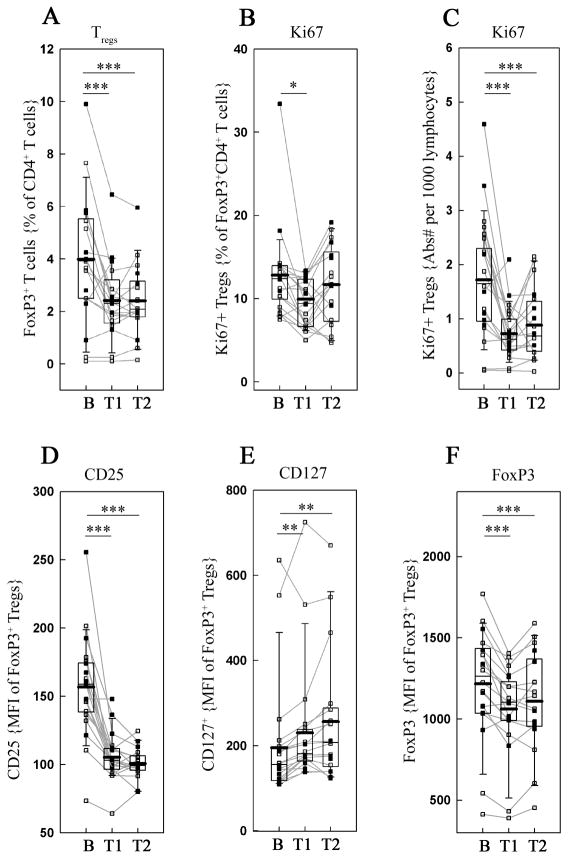
Daclizumab (Dac), an Ab against the IL-2R alpha-chain, inhibits brain inflammation in patients with multiple sclerosis, while expanding CD56(bright) immunoregulatory NK cells in vivo. We hypothesized that this unexpected expansion is paradoxically IL-2 driven; caused by the increased availability of T cell-derived IL-2 for NK cell signaling. To this end, we performed ex vivo functional analyses of CD56(bright) NK cells and T cells from patients in clinical trials with Dac. We developed in vitro models to investigate mechanisms for ex vivo observations. We observed that Dac treatment caused decreased numbers and proliferation of FoxP3(+) T regulatory cells (Tregs), a model T cell population known to be dependent on IL-2 for proliferation and survival. As anticipated, Dac therapy inhibited IL-2 signaling in all T cells; however, we also observed functional adaptation of T cells to low IL-2 signal in vivo, characterized by the concomitant enhancement of IL-7 signaling on all T cells and parallel increase of CD127 expression by Tregs. In contrast, IL-2 signaling on CD56(bright) NK cells was not inhibited by Dac and their in vivo proliferation and cytotoxicity actually increased. Mechanistic studies indicated that the activation of CD56(bright) NK cells was likely IL-2 driven, as low doses of IL-2, but not IL-15, mimicked this activation in vitro. Our study provides insight into the role that IL-2 and CD25 play in functional regulation of two important immunoregulatory cell populations in humans: FoxP3(+) Tregs and CD56(bright) NK cells.
Figures





References
-
- Cooper MA, Fehniger TA, Turner SC, Chen KS, Ghaheri BA, Ghayur T, Carson WE, Caligiuri MA. Human natural killer cells: a unique innate immunoregulatory role for the CD56 (bright) subset. Blood. 2001;97:3146–3151. - PubMed
-
- Bratke K, Kuepper M, Bade B, Virchow JC, Jr, Luttmann W. Differential expression of human granzymes A, B, and K in natural killer cells and during CD8+ T cell differentiation in peripheral blood. Eur J Immunol. 2005;35:2608–2616. - PubMed
-
- Della Chiesa M, Vitale M, Carlomagno S, Ferlazzo G, Moretta L, Moretta A. The natural killer cell-mediated killing of autologous dendritic cells is confined to a cell subset expressing CD94/NKG2A, but lacking inhibitory killer Ig-like receptors. Eur J Immunol. 2003;33:1657–1666. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
