

Regulation of dynamic polarity switching in bacteria by a Ras-like G-protein and its cognate GAP
- PMID: 20543819
- PMCID: PMC2910265
- DOI: 10.1038/emboj.2010.114
Regulation of dynamic polarity switching in bacteria by a Ras-like G-protein and its cognate GAP
Abstract
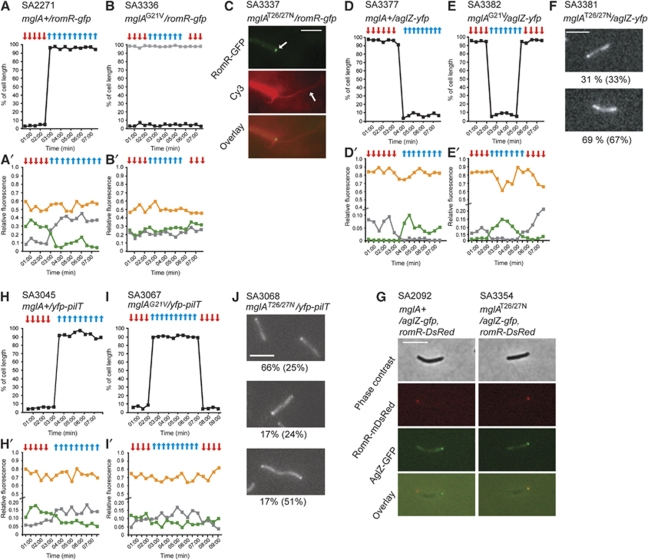
The rod-shaped cells of the bacterium Myxococcus xanthus move uni-directionally and occasionally undergo reversals during which the leading/lagging polarity axis is inverted. Cellular reversals depend on pole-to-pole relocation of motility proteins that localize to the cell poles between reversals. We show that MglA is a Ras-like G-protein and acts as a nucleotide-dependent molecular switch to regulate motility and that MglB represents a novel GTPase-activating protein (GAP) family and is the cognate GAP of MglA. Between reversals, MglA/GTP is restricted to the leading and MglB to the lagging pole defining the leading/lagging polarity axis. For reversals, the Frz chemosensory system induces the relocation of MglA/GTP to the lagging pole causing an inversion of the leading/lagging polarity axis. MglA/GTP stimulates motility by establishing correct polarity of motility proteins between reversals and reversals by inducing their pole-to-pole relocation. Thus, the function of Ras-like G-proteins and their GAPs in regulating cell polarity is found not only in eukaryotes, but also conserved in bacteria.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures






Comment in
-
Cell polarity/motility in bacteria: closer to eukaryotes than expected?EMBO J. 2010 Jul 21;29(14):2258-9. doi: 10.1038/emboj.2010.144. EMBO J. 2010. PMID: 20648047 Free PMC article. No abstract available.
Similar articles
-
A response regulator interfaces between the Frz chemosensory system and the MglA/MglB GTPase/GAP module to regulate polarity in Myxococcus xanthus.PLoS Genet. 2012 Sep;8(9):e1002951. doi: 10.1371/journal.pgen.1002951. Epub 2012 Sep 13. PLoS Genet. 2012. PMID: 23028358 Free PMC article.
-
A bacterial Ras-like small GTP-binding protein and its cognate GAP establish a dynamic spatial polarity axis to control directed motility.PLoS Biol. 2010 Jul 20;8(7):e1000430. doi: 10.1371/journal.pbio.1000430. PLoS Biol. 2010. PMID: 20652021 Free PMC article.
-
A dynamic response regulator protein modulates G-protein-dependent polarity in the bacterium Myxococcus xanthus.PLoS Genet. 2012;8(8):e1002872. doi: 10.1371/journal.pgen.1002872. Epub 2012 Aug 16. PLoS Genet. 2012. PMID: 22916026 Free PMC article.
-
Regulation of Cell Polarity in Motility and Cell Division in Myxococcus xanthus.Annu Rev Microbiol. 2017 Sep 8;71:61-78. doi: 10.1146/annurev-micro-102215-095415. Epub 2017 May 19. Annu Rev Microbiol. 2017. PMID: 28525300 Review.
-
Spatiotemporal regulation of switching front-rear cell polarity.Curr Opin Cell Biol. 2022 Jun;76:102076. doi: 10.1016/j.ceb.2022.102076. Epub 2022 Mar 31. Curr Opin Cell Biol. 2022. PMID: 35367928 Review.
Cited by
-
Myxococcus xanthus as a Model Organism for Peptidoglycan Assembly and Bacterial Morphogenesis.Microorganisms. 2021 Apr 24;9(5):916. doi: 10.3390/microorganisms9050916. Microorganisms. 2021. PMID: 33923279 Free PMC article. Review.
-
Motor-driven intracellular transport powers bacterial gliding motility.Proc Natl Acad Sci U S A. 2011 May 3;108(18):7559-64. doi: 10.1073/pnas.1101101108. Epub 2011 Apr 11. Proc Natl Acad Sci U S A. 2011. PMID: 21482768 Free PMC article.
-
Individual bacterial cells can use spatial sensing of chemical gradients to direct chemotaxis on surfaces.Nat Microbiol. 2024 Sep;9(9):2308-2322. doi: 10.1038/s41564-024-01729-3. Epub 2024 Sep 2. Nat Microbiol. 2024. PMID: 39227714 Free PMC article.
-
A molecular switch controls assembly of bacterial focal adhesions.Sci Adv. 2024 May 31;10(22):eadn2789. doi: 10.1126/sciadv.adn2789. Epub 2024 May 29. Sci Adv. 2024. PMID: 38809974 Free PMC article.
-
Uncovering the mystery of gliding motility in the myxobacteria.Annu Rev Genet. 2011;45:21-39. doi: 10.1146/annurev-genet-110410-132547. Epub 2011 Sep 9. Annu Rev Genet. 2011. PMID: 21910630 Free PMC article. Review.
References
-
- Ahmadian MR, Stege P, Scheffzek K, Wittinghofer A (1997) Confirmation of the arginine-finger hypothesis for the GAP-stimulated GTP-hydrolysis reaction of Ras. Nat Struct Biol 4: 686–689 - PubMed
-
- Bos JL, Rehmann H, Wittinghofer A (2007) GEFs and GAPs: critical elements in the control of small G proteins. Cell 129: 865–877 - PubMed
-
- Bourne HR, Sanders DA, McCormick F (1991) The GTPase superfamily: conserved structure and molecular mechanism. Nature 349: 117–127 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
