

Mechanisms used for genomic proliferation by thermophilic group II introns
- PMID: 20543989
- PMCID: PMC2882425
- DOI: 10.1371/journal.pbio.1000391
Mechanisms used for genomic proliferation by thermophilic group II introns
Abstract
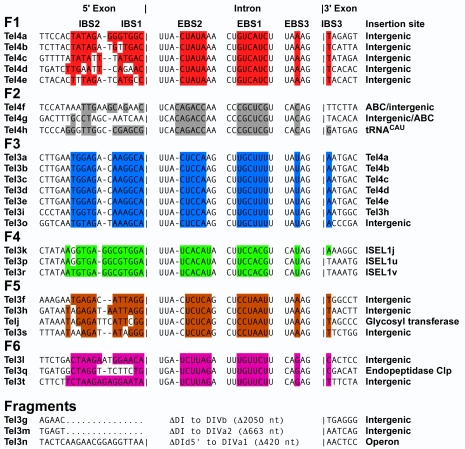
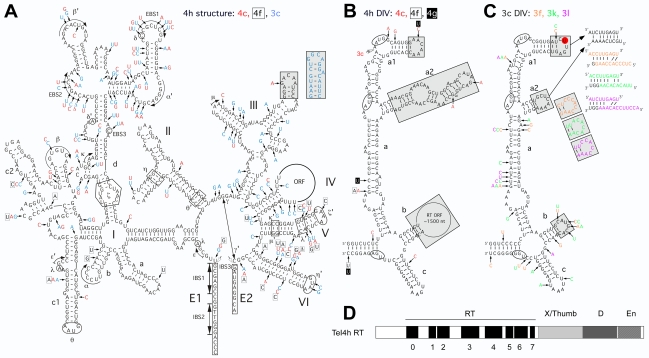
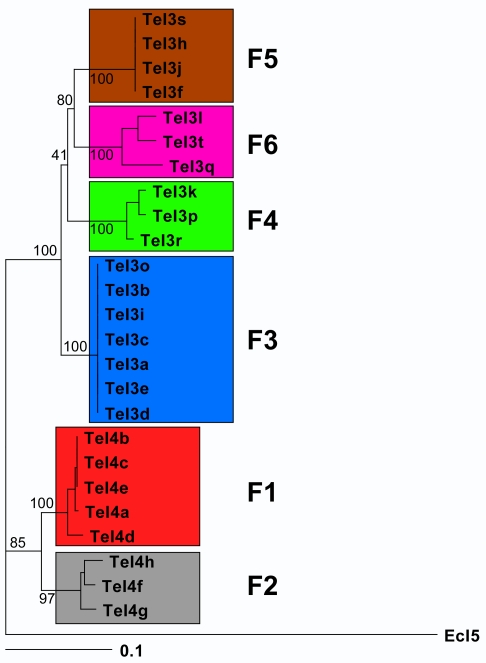
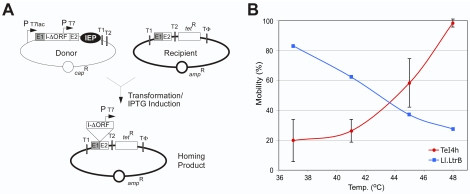
Mobile group II introns, which are found in bacterial and organellar genomes, are site-specific retroelements hypothesized to be evolutionary ancestors of spliceosomal introns and retrotransposons in higher organisms. Most bacteria, however, contain no more than one or a few group II introns, making it unclear how introns could have proliferated to higher copy numbers in eukaryotic genomes. An exception is the thermophilic cyanobacterium Thermosynechococcus elongatus, which contains 28 closely related copies of a group II intron, constituting approximately 1.3% of the genome. Here, by using a combination of bioinformatics and mobility assays at different temperatures, we identified mechanisms that contribute to the proliferation of T. elongatus group II introns. These mechanisms include divergence of DNA target specificity to avoid target site saturation; adaptation of some intron-encoded reverse transcriptases to splice and mobilize multiple degenerate introns that do not encode reverse transcriptases, leading to a common splicing apparatus; and preferential insertion within other mobile introns or insertion elements, which provide new unoccupied sites in expanding non-essential DNA regions. Additionally, unlike mesophilic group II introns, the thermophilic T. elongatus introns rely on elevated temperatures to help promote DNA strand separation, enabling access to a larger number of DNA target sites by base pairing of the intron RNA, with minimal constraint from the reverse transcriptase. Our results provide insight into group II intron proliferation mechanisms and show that higher temperatures, which are thought to have prevailed on Earth during the emergence of eukaryotes, favor intron proliferation by increasing the accessibility of DNA target sites. We also identify actively mobile thermophilic introns, which may be useful for structural studies, gene targeting in thermophiles, and as a source of thermostable reverse transcriptases.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Comment in
-
For group II introns, more heat means more mobility.PLoS Biol. 2010 Jun 8;8(6):e1000392. doi: 10.1371/journal.pbio.1000392. PLoS Biol. 2010. PMID: 20543990 Free PMC article. No abstract available.
References
-
- Lambowitz A. M, Zimmerly S. Mobile group II introns. Annu Rev Genet. 2004;38:1–35. - PubMed
-
- Pyle A. M, Lambowitz A. M. Group II introns: ribozymes that splice RNA and invade DNA. In: Gesteland R. F, Cech T, Atkins J. F, editors. The RNA world, third edition. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 2006. pp. 469–505.
-
- Michel F, Ferat J. L. Structure and activities of group II introns. Annu Rev Biochem. 1995;64:435–461. - PubMed
-
- Peebles C. L, Perlman P. S, Mecklenburg K. L, Petrillo M. L, Tabor J. H, et al. A self-splicing RNA excises an intron lariat. Cell. 1986;44:213–223. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
